Histones are the main constituents of the protein part of chromosomes of eukaryotic cells. They are rich in the amino acids arginine and lysine and have been greatly conserved during evolution. Histones pack the DNA into tight masses of chromatin. Two core histones of each class H2A, H2B, H3 and H4 assemble and are wrapped by 146 base pairs of DNA to form one octameric nucleosome. Histone tails undergo numerous post-translational modifications, which either directly or indirectly alter chromatin structure to facilitate transcriptional activation or repression or other nuclear processes. In addition to the genetic code, combinations of the different histone modifications reveal the so-called “histone code”. Histone methylation and demethylation is dynamically regulated by respectively histone methyl transferases and histone demethylases. Levels of H3K27 dimethylation are higher in silent genes than in active genes suggesting that this histone modification is associated with transcriptional repression.
H3K27me2 polyclonal antibody

Polyclonal antibody raised in rabbit against the histone H3, dimethylated at lysine 27 (H3K27me2), using a KLH-conjugated synthetic peptide.
| Lot | A1968-0024P |
|---|---|
| Concentration | 2.63 µg/µl |
| Species reactivity | Human, zebrafish |
| Type | Polyclonal |
| Purity | Affinity purified |
| Host | Rabbit |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP * | 2 μg/ChIP | Fig 1 |
| ELISA | 1:1,000 | Fig 2 |
| Dot Blotting | 1:50,000 | Fig 3 |
| Western Blotting | 1:1,000 | Fig 4 |
| Immunofluorescence | 1:500 | Fig 5 |
- Validation Data
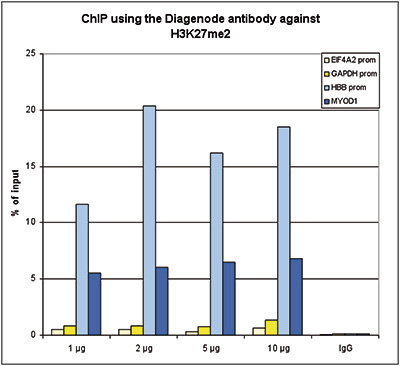
Figure 1. ChIP results obtained with the Diagenode antibody directed against H3K27me2
ChIP assays were performed using human HeLa cells, the Diagenode antibody against H3K27me2 (cat. No. C15410046) and optimized PCR primer sets for qPCR. ChIP was performed with the “Auto Histone ChIP-seq” kit (cat. No. AB-Auto02-A100) on the SX-8G IP-Star Compact automated system, using sheared chromatin from 1 million cells. A titration of the antibody consisting of 1, 2, 5, and 10 μg per ChIP experiment was analysed. IgG (2 μg/IP) was used as negative IP control. QPCR was performed with primers for the promoter of the active GAPDH and EIF4A2 genes, used as negative controls, and for the promoter of the inactive HBB and the coding region of the inactive MYOD1 genes, used as positive controls. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis). These results are in accordance with the observation that H3K27me2 is preferably present at silent genes.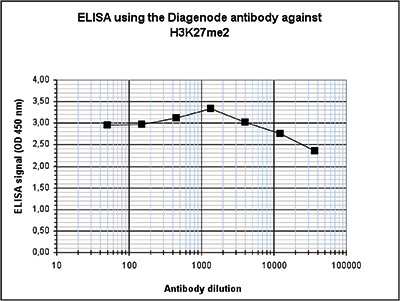
Figure 2. Determination of the antibody titer
To determine the titer of the antibody, an ELISA was performed using a serial dilution of the Diagenode antibody directed against H3K27me2 (cat. No. C15410046). The antigen used was a peptide containing the histone modification of interest. By plotting the absorbance against the antibody dilution (Figure 2), the titer of the antibody was estimated to be 1:480,000.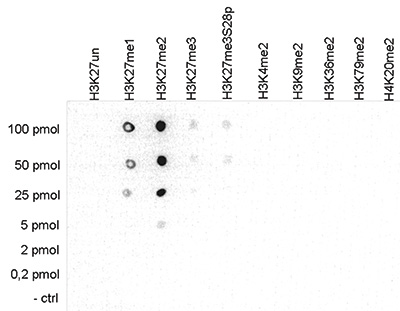
Figure 3. Cross reactivity tests using the Diagenode antibody directed against H3K27me2
A Dot Blot analysis was performed to test the cross reactivity of the Diagenode antibody against H3K27me2 (cat. No. C15410046) with peptides containing other modifications of histone H3 and H4 and the unmodified sequence. One hundred to 0.2 pmol of peptide containing the respective histone modification were spotted on a membrane. The antibody was used at a dilution of 1:50,000. Figure 3 shows a high specificity of the antibody for the modification of interest.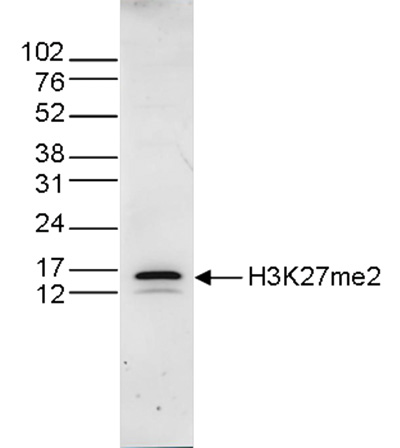
Figure 4. Western blot analysis using the Diagenode antibody directed against H3K27me2
Histone extracts (15 μg) from HeLa cells were analysed by Western blot using the Diagenode antibody against H3K27me2 (cat. No. C15410046) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left.
Figure 5. Immunofluorescence using the Diagenode antibody directed against H3K27me2
HeLa cells were stained with the Diagenode antibody against H3K27me2 (cat. No. C15410046) and with DAPI. Cells were fixed with 4% formaldehyde for 10’ and blocked with PBS/TX-100 containing 5% normal goat serum and 1% BSA. The cells were immunofluorescently labelled with the H3K27me2 antibody (left) diluted 1:500 in blocking solution followed by an anti-rabbit antibody conjugated to Alexa488. The middle panel shows staining of the nuclei with DAPI. A merge of the two stainings is shown on the right. - 出版物

How to properly cite our product/service in your work
We strongly recommend using this: H3K27me2 polyclonal antibody (Hologic Diagenode Cat# C15410046 Lot# A1968-0024P). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
Motif distribution and DNA methylation underlie distinct Cdx2 binding during development and homeostasis
Alireza Lorzadeh et al.
Transcription factors guide tissue development by binding to developmental stage-specific targets and establishing an appropriate enhancer landscape. In turn, DNA and chromatin modifications direct the genomic binding of transcription factors. However, how transcription factors navigate chromatin features to selecti...KDM6B drives epigenetic reprogramming associated with lymphoid stromal cell early commitment and immune properties
Sylvestre M. et al.
Mature lymphoid stromal cells (LSCs) are key organizers of immune responses within secondary lymphoid organs. Similarly, inflammation-driven tertiary lymphoid structures depend on immunofibroblasts producing lymphoid cytokines and chemokines. Recent studies have explored the origin and heterogeneity of LSC/immunofib...Nox4 promotes endothelial differentiation through chromatin remodeling.
Hahner F. et al.
RATIONALE: Nox4 is a constitutively active NADPH oxidase that constantly produces low levels of HO. Thereby, Nox4 contributes to cell homeostasis and long-term processes, such as differentiation. The high expression of Nox4 seen in endothelial cells contrasts with the low abundance of Nox4 in stem cells, which are a...Forskolin Sensitizes Human Acute Myeloid Leukemia Cells to H3K27me2/3 Demethylases GSKJ4 Inhibitor via Protein Kinase A.
Illiano M, Conte M, Sapio L, Nebbioso A, Spina A, Altucci L, Naviglio S
Acute myeloid leukemia (AML) is an aggressive hematological malignancy occurring very often in older adults, with poor prognosis depending on both rapid disease progression and drug resistance occurrence. Therefore, new therapeutic approaches are demanded. Epigenetic marks play a relevant role in AML. GSKJ4 is a nov...The Polycomb-Dependent Epigenome Controls β Cell Dysfunction, Dedifferentiation, and Diabetes.
Lu TT, Heyne S, Dror E, Casas E, Leonhardt L, Boenke T, Yang CH, Sagar , Arrigoni L, Dalgaard K, Teperino R, Enders L, Selvaraj M, Ruf M, Raja SJ, Xie H, Boenisch U, Orkin SH, Lynn FC, Hoffman BG, Grün D, Vavouri T, Lempradl AM, Pospisilik JA
To date, it remains largely unclear to what extent chromatin machinery contributes to the susceptibility and progression of complex diseases. Here, we combine deep epigenome mapping with single-cell transcriptomics to mine for evidence of chromatin dysregulation in type 2 diabetes. We find two chromatin-state signat...Role of Annexin gene and its regulation during zebrafish caudal fin regeneration
Saxena S, Purushothaman S, Meghah V, Bhatti B, Poruri A, Meena Lakshmi MG, Sarath Babu N, Murthy CL, Mandal KK, Kumar A, Idris MM
The molecular mechanism of epimorphic regeneration is elusive due to its complexity and limitation in mammals. Epigenetic regulatory mechanisms play a crucial role in development and regeneration. This investigation attempted to reveal the role of epigenetic regulatory mechanisms, such as histone H3 and H4 lysine ac...