Histones are the main constituents of the protein part of chromosomes of eukaryotic cells. They are rich in the amino acids arginine and lysine and have been greatly conserved during evolution. Histones pack the DNA into tight masses of chromatin. Two core histones of each class H2A, H2B, H3 and H4 assemble and are wrapped by 146 base pairs of DNA to form one octameric nucleosome. Histone tails undergo numerous post-translational modifications, which either directly or indirectly alter chromatin structure to facilitate transcriptional activation or repression or other nuclear processes. In addition to the genetic code, combinations of the different histone modifications reveal the so-called “histone code”. Histone methylation and demethylation is dynamically regulated by respectively histone methyl transferases and histone demethylases.
H3K27me3 Antibody (sample size)

Monoclonal antibody raised in mouse against histone H3 trimethylated at lysine 27 (H3K27me3), using a KLH-conjugated synthetic peptide.
| Lot | 001-16 |
|---|---|
| Concentration | 1 µg/µl |
| Species reactivity | Human, Nematodes, Magnaporthe oryzae: positive. Other species: not tested. |
| Type | Monoclonal |
| Purity | Protein A purified monoclonal antibody. |
| Host | Mouse |
| Storage Conditions | Store at -20°C; for long storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Storage Buffer | PBS containing 0.05% azide. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP * | 1-2 µg/ChIP | Fig 1 |
| CUT&TAG | 1 µg | Fig 2 |
| ELISA | 1:3,000 | Fig 3 |
| Western Blotting | 1:1,000 | Fig 4 |
| Immunofluorescence | 1:500 | Fig 5 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-5 µg per IP.
- Validation Data
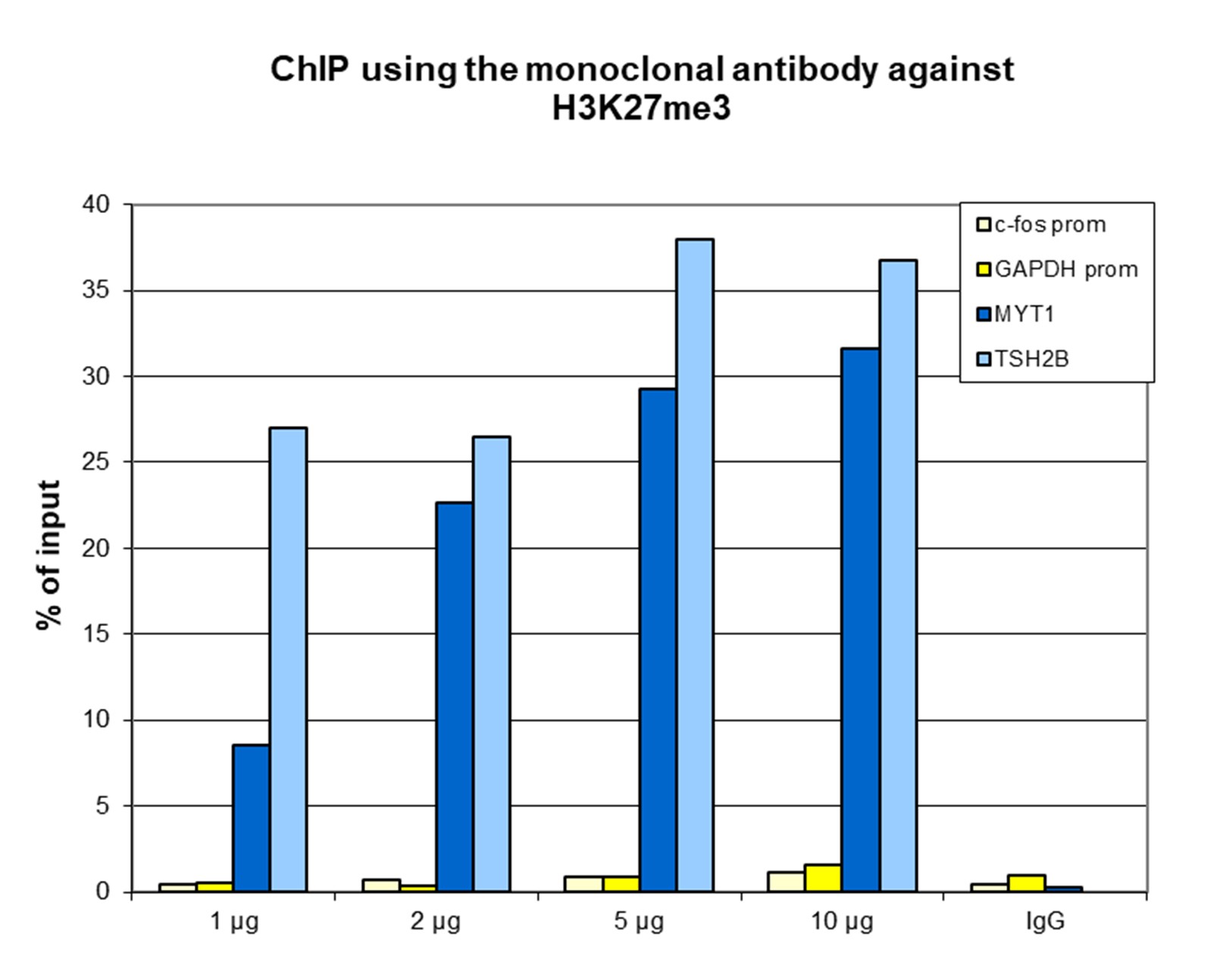
Figure 1. ChIP results obtained with the monoclonal antibody directed against H3K27me3
ChIP assays were performed using human HeLa cells, the monoclonal antibody against H3K27me3 (cat. No. C15200181) and optimized PCR primer sets for qPCR. ChIP was performed with the “iDeal ChIP-seq” kit (cat. No. C01010051) on sheared chromatin from 1 million cells. A titration of the antibody consisting of 1, 2, 5, and 10 µg per ChIP experiment was analysed. IgG (2 µg/IP) was used as negative IP control. QPCR was performed with primers for the promoters of the active genes c-fos (cat. No. C17011004) and GAPDH as negative controls, and for the coding regions of the inactive genes MYT1 and TSH2B (cat. No. C17011041) as positive controls. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis). These results are in accordance with the observation that H3K27me3 is preferably present at inactive genes.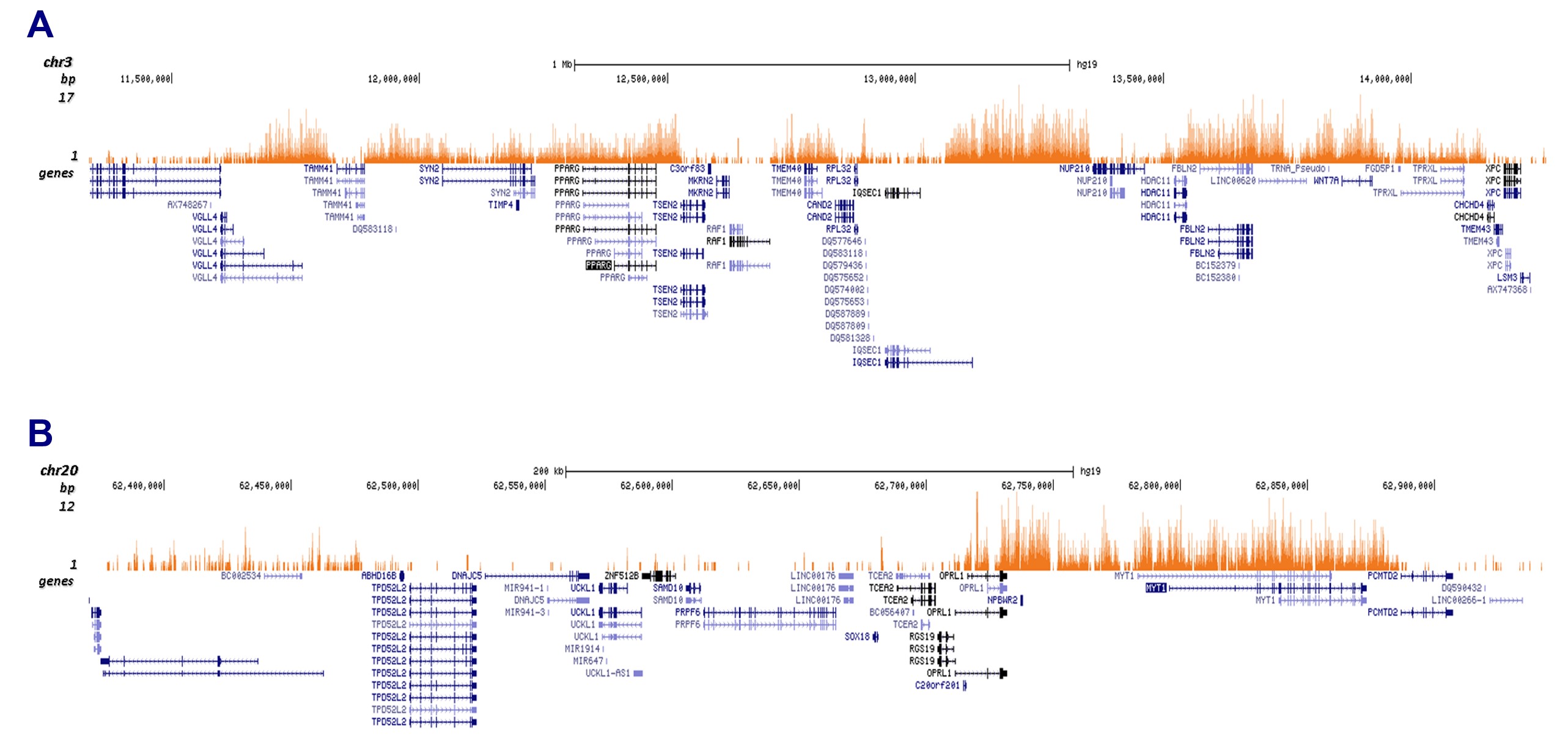
Figure 2. Cut&Tag results obtained with the monoclonal antibody directed against H3K27me3
CUT&TAG (Kaya-Okur, H.S., Nat Commun 10, 1930, 2019) was performed on 50,000 K562 cells using 1 µg of the monoclonal antibody against H3K27me3 (cat. No. C15200181) and the Diagenode pA-Tn5 transposase (C01070001). The libraries were subsequently analysed on an Illumina NextSeq 500 sequencer (2x75 paired-end reads) according to the manufacturer’s instructions. The tags were aligned to the human genome (hg19) using the BWA algorithm. Figure 3 shows the peak distribution in 2 genomic regions on chromosome 3 and 20 (figure 2A and B, respectively).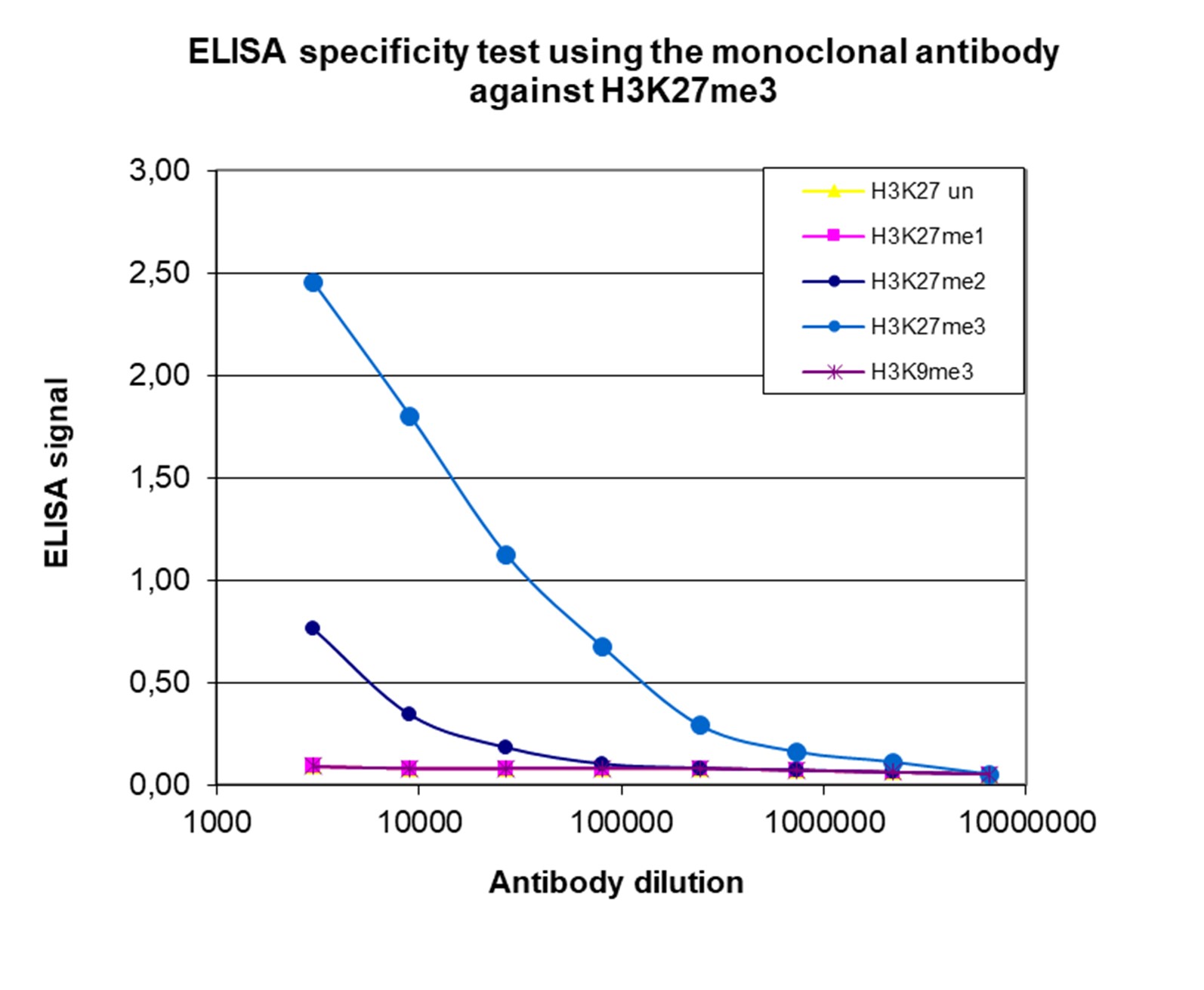
Figure 3. Cross reactivity of the monoclonal antibody directed against H3K27me3
To test the specificity an ELISA was performed using a serial dilution of the monoclonal antibody against H3K27me3 (cat. No. C15200181). The wells were coated with peptides containing the unmodified H3K27 region as well as the mono-, di- and trimethylated H3K27 and the trimethylated H3K9. Figure 3 shows a high specificity of the antibody for the peptide containing the modification of interest.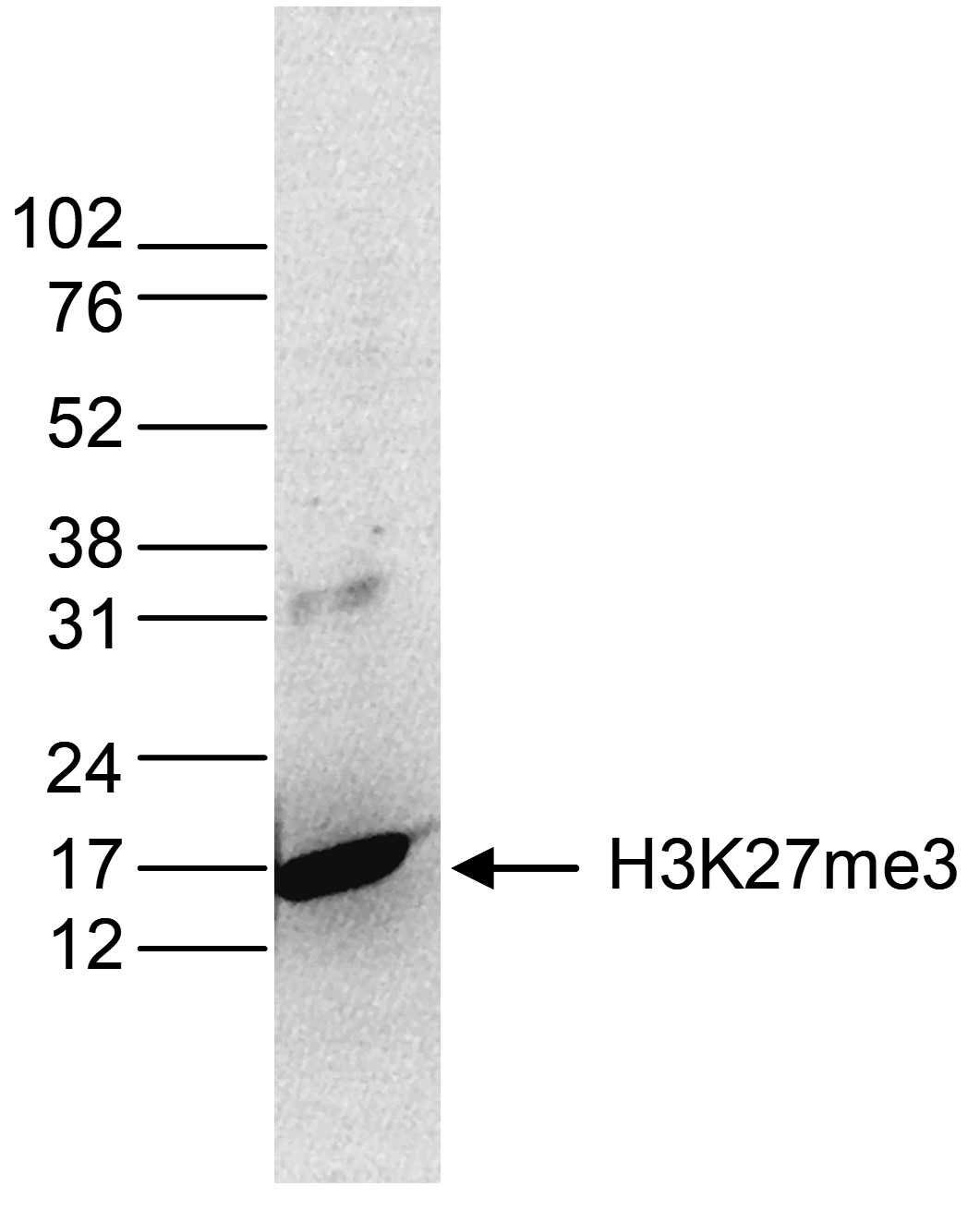
Figure 4. Western blot analysis using the monoclonal antibody directed against H3K27me3
Histone extracts (15 µg) from HeLa cells were analysed by Western blot using the monoclonal antibody against H3K27me3 (cat. No. C15200181) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left.
Figure 5. Immunofluorescence using the monoclonal antibody directed against H3K27me3
HeLa cells were stained with the antibody against H3K27me3 (cat. No. C15200181) and with DAPI. Cells were fixed with 4% formaldehyde for 10’ and blocked with PBS/TX-100 containing 5% normal goat serum and 1% BSA. The cells were immunofluorescently labelled with the H3K27me3 antibody (left) diluted 1:500 in blocking solution followed by an anti-mouse antibody conjugated to Alexa594. The middle panel shows staining of the nuclei with DAPI. A merge of the two stainings is shown on the right. - Publications

How to properly cite our product/service in your work
We strongly recommend using this: H3K27me3 Antibody (sample size) (Hologic Diagenode Cat# C15200181-10 Lot# 001-16). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
PHF19 drives the formation of PRC2 clusters to enhance motility in TNBC cells
Pelzer, Nina et al.
Polycomb repressive complex 2 (PRC2) is a key regulator of transcriptional repression and chromatin organization, essential in development and disease. While its enzymatic activity is well characterized, the factors governing PRC2 subnuclear organization, particularly in cancer cells, are largely unknown. Here, we...Constitutive expression of the transcriptional co-activator IκBζ promotes melanoma growth and immunotherapy resistance
Kolb, Antonia et al.
IκBζ, a rather unknown co-regulator of NF-κB, can either activate or repress a subset of NF-κB target genes. While its role as an inducibly expressed, transcriptional regulator of cytokines and chemokines in immune cells is established, IκBζ's function in solid cancer remains un...Chromatin profiling reveals TFAP4 as a critical transcriptional regulator of bovine satellite cell differentiation
Pengcheng Lyu et al.
Background Satellite cells are myogenic precursor cells in adult skeletal muscle and play a crucial role in skeletal muscle regeneration, maintenance, and growth. Like embryonic myoblasts, satellite cells have the ability to proliferate, differentiate, and fuse to form multinucleated myofibers. In this study, we ai...Tracing the emergence of primordial germ cells from bilaminar disc rabbitembryos and pluripotent stem cells.
Kobayashi Toshihiro et al.
Rabbit embryos develop as bilaminar discs at gastrulation as in humans and most other mammals, whereas rodents develop as egg cylinders. Primordial germ cells (PGCs) appear to originate during gastrulation according to many systematic studies on mammalian embryos. Here, we show that rabbit PGC (rbPGC) specification ...Histone modification dynamics at H3K27 are associated with alteredtranscription of in planta induced genes in Magnaporthe oryzae.
Zhang, Wei and Huang, Jun and Cook, David E
Transcriptional dynamic in response to environmental and developmental cues are fundamental to biology, yet many mechanistic aspects are poorly understood. One such example is fungal plant pathogens, which use secreted proteins and small molecules, termed effectors, to suppress host immunity and promote colonization...Intergenerationally Maintained Histone H4 Lysine 16 Acetylation Is Instructive for Future Gene Activation.
Samata M, Alexiadis A, Richard G, Georgiev P, Nuebler J, Kulkarni T, Renschler G, Basilicata MF, Zenk FL, Shvedunova M, Semplicio G, Mirny L, Iovino N, Akhtar A
Before zygotic genome activation (ZGA), the quiescent genome undergoes reprogramming to transition into the transcriptionally active state. However, the mechanisms underlying euchromatin establishment during early embryogenesis remain poorly understood. Here, we show that histone H4 lysine 16 acetylation (H4K16ac) i...Pro-inflammatory cytokines activate hypoxia-inducible factor 3α via epigenetic changes in mesenchymal stromal/stem cells.
Cuomo F, Coppola A, Botti C, Maione C, Forte A, Scisciola L, Liguori G, Caiafa I, Ursini MV, Galderisi U, Cipollaro M, Altucci L, Cobellis G
Human mesenchymal stromal/stem cells (hMSCs) emerged as a promising therapeutic tool for ischemic disorders, due to their ability to regenerate damaged tissues, promote angiogenesis and reduce inflammation, leading to encouraging, but still limited results. The outcomes in clinical trials exploring hMSC therapy are ...Chemical Biology Approaches for Characterization of Epigenetic Regulators
Barsyte-Lovejoy D, Szewczyk MM, Prinos P, Lima-Fernandes E, Ackloo S, Arrowsmith CH
Chemical biology approaches are a powerful means to functionally characterize epigenetic regulators such as histone modifying enzymes. We outline experimental protocols and best practices for the cellular characterization and use of “chemical probes” that selectively inhibit protein methyltransferases, m...Germline organization in Strongyloides nematodes reveals alternative differentiation and regulation mechanisms.
Kulkarni A et al.
Nematodes of the genus Strongyloides are important parasites of vertebrates including man. Currently, little is known about their germline organization or reproductive biology and how this influences their parasitic life strategies. Here, we analyze the structure of the germline in several Strongyloides and closely ...An Orally Bioavailable Chemical Probe of the Lysine Methyltransferases EZH2 and EZH1.
Konze KD, Ma A, Li F, Barsyte-Lovejoy D, Parton T, Macnevin CJ, Liu F, Gao C, Huang XP, Kuznetsova E, Rougie M, Jiang A, Pattenden SG, Norris JL, James LI, Roth BL, Brown PJ, Frye SV, Arrowsmith CH, Hahn KM, Wang GG, Vedadi M, Jin J
EZH2 or EZH1 is the catalytic subunit of the polycomb repressive complex 2 that catalyzes methylation of histone H3 lysine 27 (H3K27). The trimethylation of H3K27 (H3K27me3) is a transcriptionally repressive post-translational modification. Overexpression of EZH2 and hypertrimethylation of H3K27 have been implicated...