Histones are the main constituents of the protein part of chromosomes of eukaryotic cells. They are rich in the amino acids arginine and lysine and have been greatly conserved during evolution. Histones pack the DNA into tight masses of chromatin. Two core histones of each class H2A, H2B, H3 and H4 assemble and are wrapped by 146 base pairs of DNA to form one octameric nucleosome. Histone tails undergo numerous post-translational modifications, which either directly or indirectly alter chromatin structure to facilitate transcriptional activation or repression or other nuclear processes. In addition to the genetic code, combinations of the different histone modifications reveal the so-called “histone code”. Histone methylation and demethylation is dynamically regulated by respectively histone methyl transferases and histone demethylases. Trimethylation of histone H3 on K79 was shown to be more present at active promotors than at silent promotors.
H3K79me3 Antibody

Polyclonal antibody raised in rabbit against histone H3 containing the trimethylated lysine 79 (H3K79me3), using a KLH-conjugated synthetic peptide.
| Lot | A2014P |
|---|---|
| Concentration | 0.53 µg/µl |
| Species reactivity | Human, mouse, yeast: positive. Other species: not tested. |
| Type | Polyclonal |
| Purity | Affinity purified polyclonal antibody |
| Host | Rabbit |
| Storage Conditions | Store at -20°C; for long storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Storage Buffer | PBS containing 0.05% azide and 0.05% ProClin 300. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 1-2 μg/ChIP | Fig 1, 2 |
| ELISA | 1:500 – 1/1,000 | Fig 3 |
| Dot Blotting | 1:20,000 | Fig 4 |
| Western Blotting | 1:1,00 | Fig 5 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 0.5-5 μg per IP.
- Validation Data
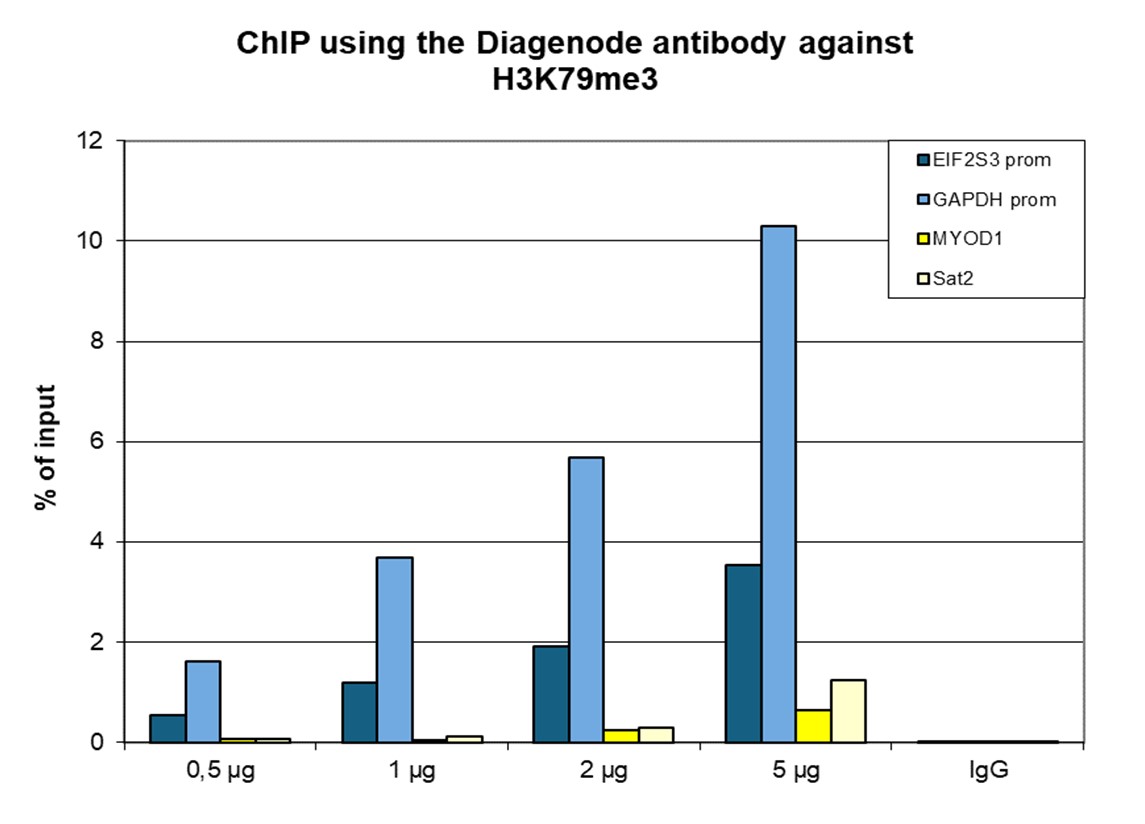
Figure 1. ChIP results obtained with the Diagenode antibody directed against H3K79me3
ChIP was performed with the Diagenode antibody against H3K79me3 (cat. No. C15410068) on sheared chromatin from 1 million HeLaS3 cells using the “Auto Histone ChIP-seq” kit and the SX-8G IP-Star automated system. A titration of the antibody consisting of 1, 2, 5 and 10 µg per ChIP experiment was analysed. IgG (5 µg/IP) was used as negative IP control. Quantitative PCR was performed with primers for the GAPDH promoter and for exon 2 of the inactive myoglobin gene. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis). These results are in accordance with the observation that H3K79me3 shows a preference for active promoters.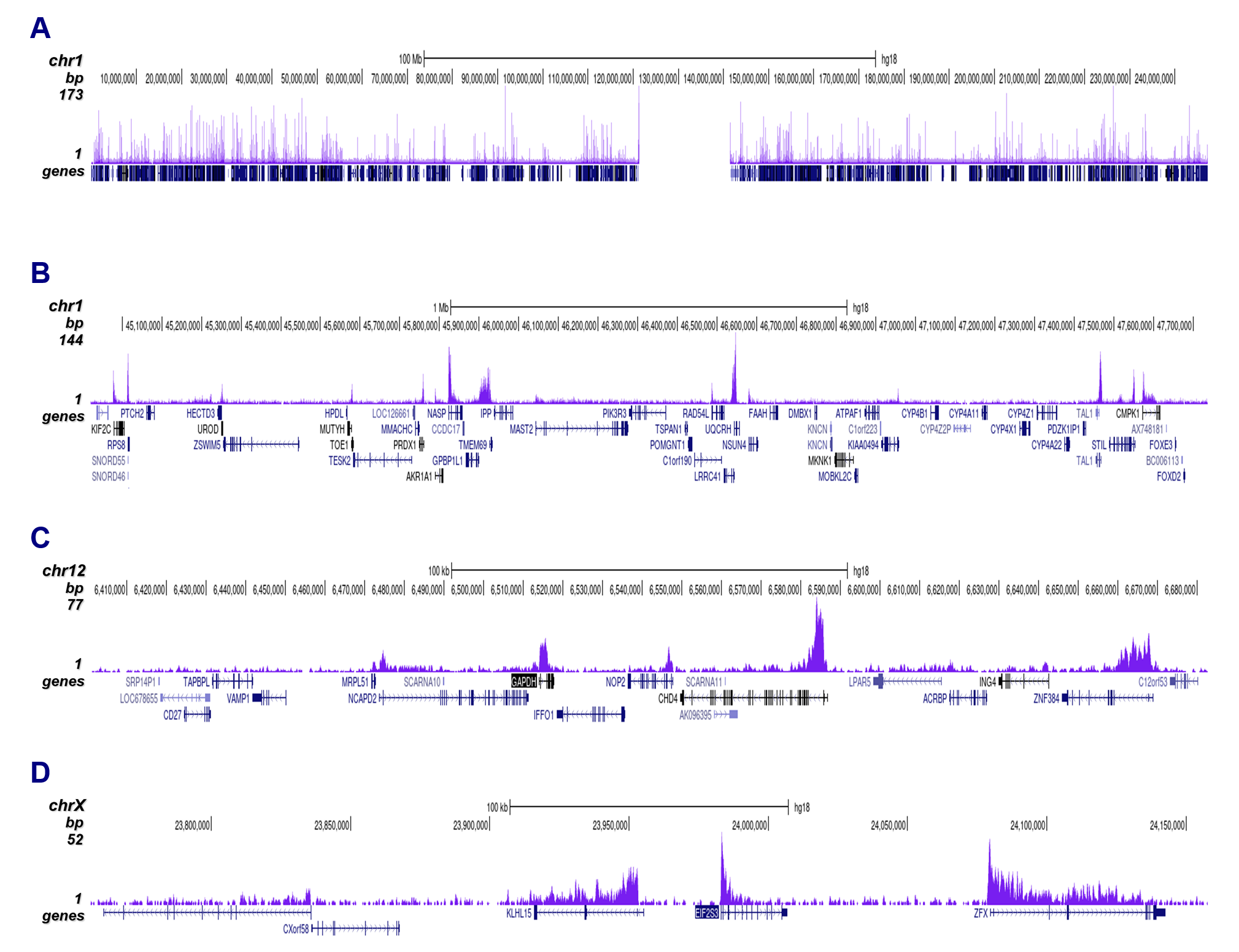
Figure 2. ChIP-seq results obtained with the Diagenode antibody directed against H3K79me3
ChIP was performed on 100,000 K562 cells using 1 µg of the Diagenode antibody against H3K79me3 (cat. No. C15410068). The IP'd DNA was analysed on an Illumina Genome Analyzer. Library preparation, cluster generation and sequencing were performed according to the manufacturer's instructions. The 36 bp tags were aligned to the human genome using the ELAND algorithm. Figure 2 shows the peak distribution along the complete sequence and a 3 Mb region of human chromosome 1 (figure 2A and B), in a 2 genomic regions surrounding the GAPDH and EIF2S3 positive control genes (figure 2C and D).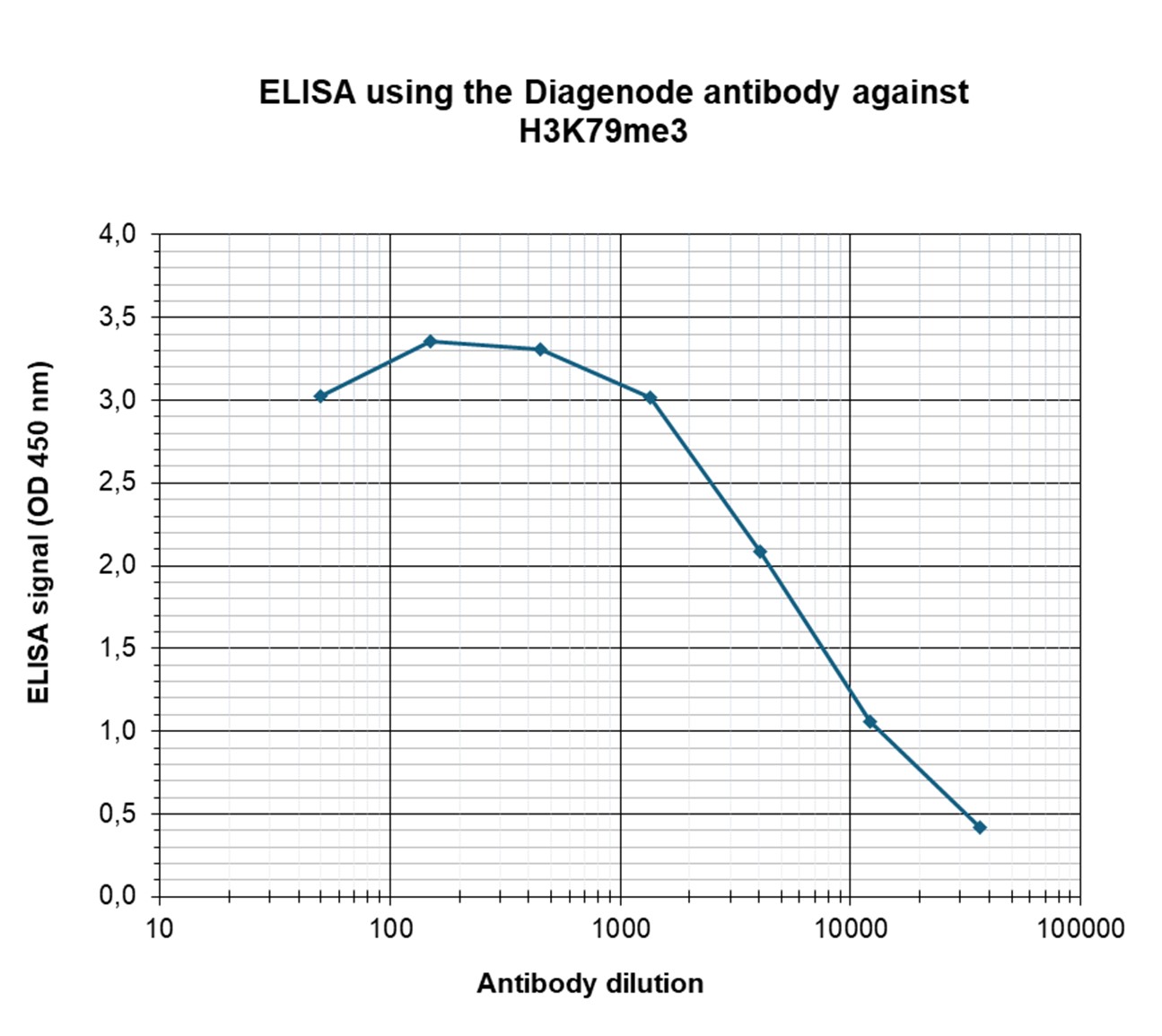
Figure 3. Determination of the antibody titer
To determine the titer of the antibody, an ELISA was performed using a serial dilution of Diagenode antibody directed against H3K79me3 (cat. No. C15410068) in antigen coated wells. The antigen used was a peptide containing the histone modification of interest. By plotting the absorbance against the antibody dilution (Figure 3), the titer of the antibody was estimated to be 1:8,500.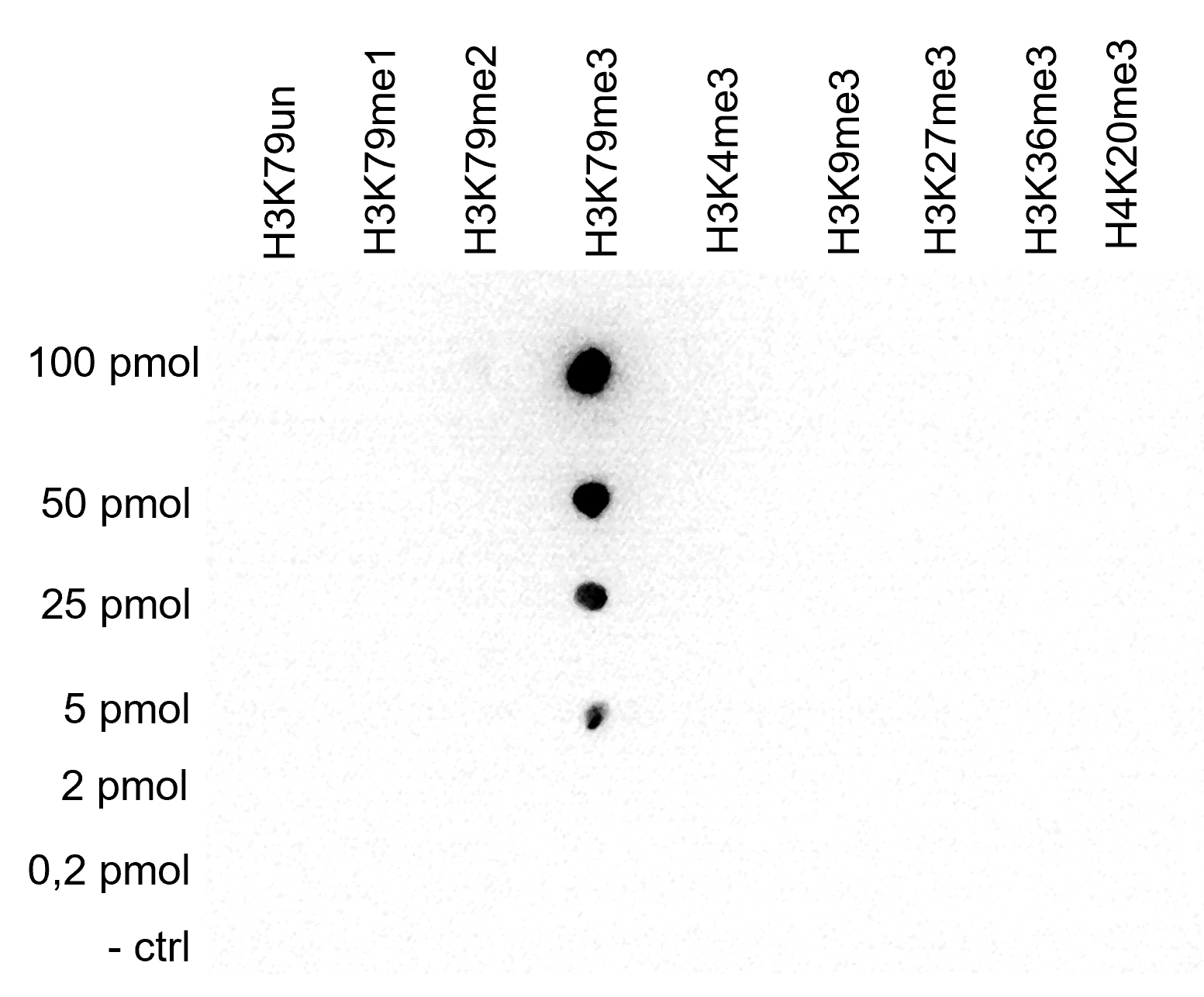
Figure 4. Cross reactivity tests using the Diagenode antibody directed against H3K79me3
A Dot Blot analysis was performed to test the cross reactivity of the Diagenode antibody against H3K79me3 (cat. No.C15410068) with peptides containing other histone modifications and the unmodified H3K79. One hundred to 0.2 pmol of the respective peptides were spotted on a membrane. The antibody was used at a dilution of 1:100,000. Figure 4 shows a high specificity of the antibody for the modification of interest.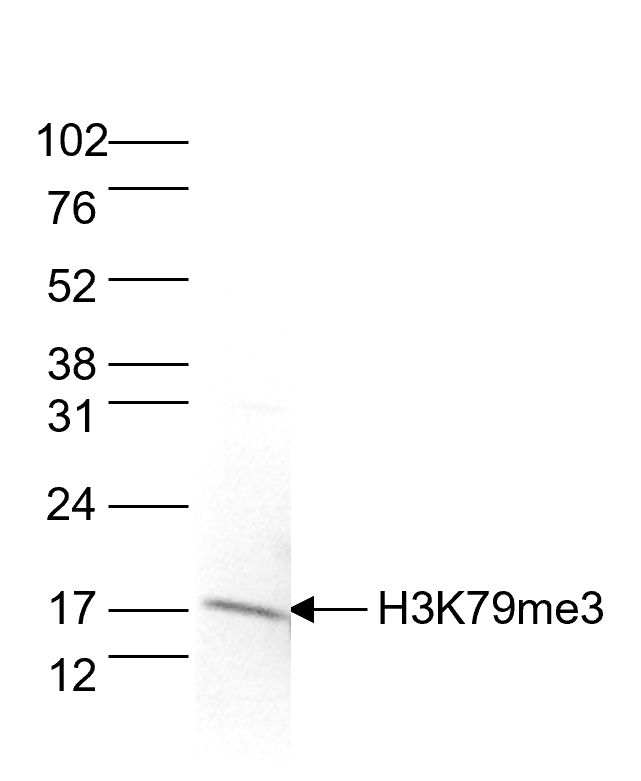
Figure 5. Western blot analysis using the Diagenode antibody directed against H3K79me3
Histone extracts of HeLa cells (15 µg) were analysed by Western blot using the Diagenode antibody against H3K79me3 (cat. No.C15410068), diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The molecular weight marker is shown on the right; the location of the protein of interest is indicated on the left. - Publications

How to properly cite our product/service in your work
We strongly recommend using this: H3K79me3 Antibody (Hologic Diagenode Cat# C15410068 Lot# A2014P). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
Distinct silencer states generate epigenetic states of heterochromatin.
Saxton Daniel S and Rine Jasper
Heterochromatic loci can exhibit different transcriptional states in genetically identical cells. A popular model posits that the inheritance of modified histones is sufficient for inheritance of the silenced state. However, silencing inheritance requires silencers and therefore cannot be driven by the inheritance o...DOT1L O-GlcNAcylation promotes its protein stability andMLL-fusion leukemia cell proliferation.
Song Tanjing et al.
Histone lysine methylation functions at the interface of the extracellular environment and intracellular gene expression. DOT1L is a versatile histone H3K79 methyltransferase with a prominent role in MLL-fusion leukemia, yet little is known about how DOT1L responds to extracellular stimuli. Here, we report that DOT1...The histone methyltransferase DOT1L is required for proper DNA damage response, DNA repair, and modulates chemotherapy responsiveness.
Kari V, Raul SK, Henck JM, Kitz J, Kramer F, Kosinsky RL, Übelmesser N, Mansour WY, Eggert J, Spitzner M, Najafova Z, Bastians H, Grade M, Gaedcke J, Wegwitz F, Johnsen SA
BACKGROUND: Disruptor of telomeric silencing 1-like (DOT1L) is a non-SET domain containing methyltransferase known to catalyze mono-, di-, and tri-methylation of histone 3 on lysine 79 (H3K79me). DOT1L-mediated H3K79me has been implicated in chromatin-associated functions including gene transcription, heterochromati...Tri-methylation of H3K79 is decreased in TGF-β1-induced epithelial-to-mesenchymal transition in lung cancer
Evanno E. et al.
BACKGROUND: The epithelial-to-mesenchymal transition (EMT) enables epithelial cancer cells to acquire mesenchymal features and contributes to metastasis and resistance to treatment. This process involves epigenetic reprogramming for gene expression. We explored global histone modifications during TGF-β1-indu...DOT1L Activity Promotes Proliferation and Protects Cortical Neural Stem Cells from Activation of ATF4-DDIT3-Mediated ER Stress In Vitro
Roidl D, Hellbach N, Bovio PP, Villarreal A, Heidrich S, Nestel S, Grüning BA, Boenisch U, Vogel T
Growing evidence suggests that the lysine methyltransferase DOT1L/KMT4 has important roles in proliferation, survival, and differentiation of stem cells in development and in disease. We investigated the function of DOT1L in neural stem cells (NSCs) of the cerebral cortex. The pharmacological inhibition and shRNA-me...MLL-Rearranged Acute Lymphoblastic Leukemias Activate BCL-2 through H3K79 Methylation and Are Sensitive to the BCL-2-Specific Antagonist ABT-199
Benito JM et al.
Targeted therapies designed to exploit specific molecular pathways in aggressive cancers are an exciting area of current research. Mixed Lineage Leukemia (MLL) mutations such as the t(4;11) translocation cause aggressive leukemias that are refractory to conventional treatment. The t(4;11) translocation produces an M...Degree of recruitment of DOT1L to MLL-AF9 defines level of H3K79 Di- and tri-methylation on target genes and transformation potential.
Kuntimaddi A, Achille NJ, Thorpe J, Lokken AA, Singh R, Hemenway CS, Adli M, Zeleznik-Le NJ, Bushweller JH
The MLL gene is a common target of chromosomal translocations found in human leukemia. MLL-fusion leukemia has a consistently poor outcome. One of the most common translocation partners is AF9 (MLLT3). MLL-AF9 recruits DOT1L, a histone 3 lysine 79 methyltransferase (H3K79me1/me2/me3), leading to aberrant gene transc...Anticheckpoint pathways at telomeres in yeast
Ribeyre Cyril, Shore David
Telomeres hide (or ‘cap’) chromosome ends from DNA-damage surveillance mechanisms that arrest the cell cycle and promote repair, but the checkpoint status of telomeres is not well understood. Here we characterize the response in Saccharomyces cerevisiae to DNA double-strand breaks (DSBs) flanked by varying amounts o...Promoter-exon relationship of H3 lysine 9, 27, 36 and 79 methylation on pluripotency-associated genes.
Barrand S, Andersen IS, Collas P
Evidence links pluripotency to a gene regulatory network organized by the transcription factors Oct4, Nanog and Sox2. Expression of these genes is controlled by epigenetic modifications on regulatory regions. However, little is known on profiles of trimethylated H3 lysine residues on coding regions of these genes in...Chromatin states of core pluripotency-associated genes in pluripotent, multipotent and differentiated cells.
Barrand S, Collas P
Oct4, Nanog and Sox2 constitute a core of transcription factors controlling pluripotency. Differentiation and reprogramming studies have unraveled a few epigenetic modifications associated in relation to the expression state of OCT4, NANOG and SOX2. There is, however, no comprehensive map of chromatin states on thes...