Histones are present in the chromosomes of eukaryotic cells. They are rich in the amino acids arginine and lysine and have been greatly conserved during evolution. Histones pack the DNA into tight masses of chromatin. Two core histones of each class H2A, H2B, H3 and H4 assemble and are wrapped by 146 base pairs of DNA to form one octameric nucleosome. Histone tails undergo numerous post-translational modifications, which either directly or indirectly alter chromatin structure to facilitate transcriptional activation or repression or other nuclear processes. In addition to the genetic code, combinations of the different histone modifications reveal the so-called “histone code”. Histone methylation and demethylation is dynamically regulated by respectively histone methyl transferases and histone demethylases. Methylation of histone H3K9 is associated with gene repression.
H3K9me3 Antibody (sample size)

Monoclonal antibody raised in mouse against histone H3 trimethylated at lysine 9 (H3K9me3), using a KLH-conjugated synthetic peptide.
| Lot | 003 |
|---|---|
| Concentration | 1.7 µg/µl |
| Species reactivity | Human, mouse, fungi: positive. Other species: not tested |
| Type | Monoclonal, ChIP-grade, ChIP-seq grade, CUT&Tag-grade |
| Purity | Protein A purified |
| Host | Mouse |
| Storage Conditions | Store at -20°C; for long storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Storage Buffer | PBS containing 0.05% azide. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 0.5 - 1 μg/ChIP | Fig 1, 2 |
| CUT&Tag | 1 µg | Fig 3 |
| ELISA | 1:100 | Fig 4 |
| Dot Blotting | 1:100,000 | Fig 5 |
| Western Blotting | 1:1,000 | Fig 6 |
| IF | 1:500 | Fig 7 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-5 μg per IP.
- Validation Data
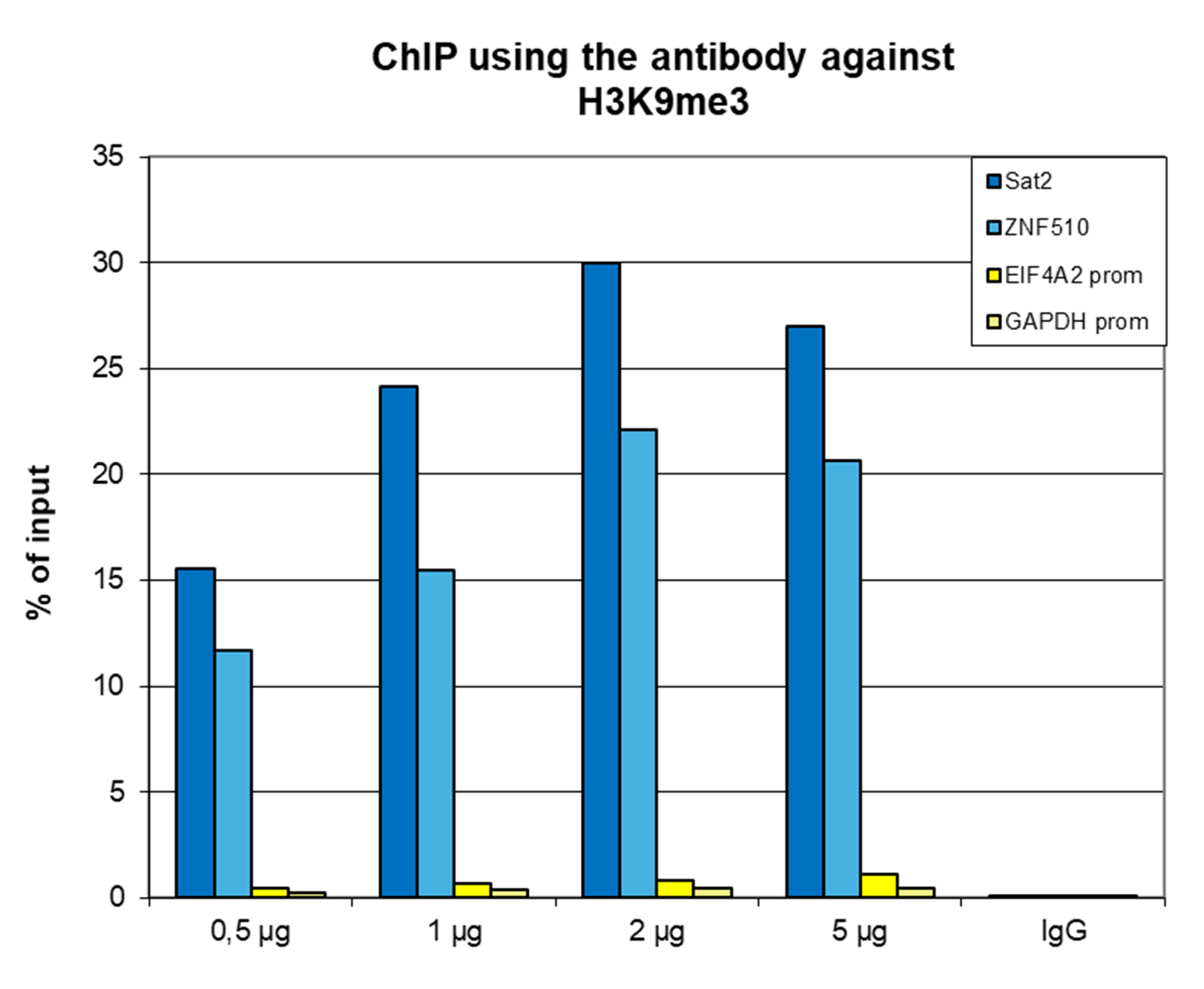
Figure 1. ChIP results obtained with the monoclonal antibody directed against H3K9me3
ChIP assays were performed on human HeLa cells using the monoclonal antibody against H3K9me3 (cat. No. C15200146). ChIP was performed with the iDeal ChIP-seq kit (cat. No. C01010051), using sheared chromatin from 500,000 cells. A titration consisting of 0.5, 1, 2 and 5 µg of antibody per ChIP experiment was analyzed. IgG (1 µg/IP) was used as a negative IP control. qPCR was performed with primers for the promoters of the active EIF4A2 and GAPDH genes (used as negative controls) and for the ZNF510 gene and the Sat2 satellite repeat region (used as positive controls). Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).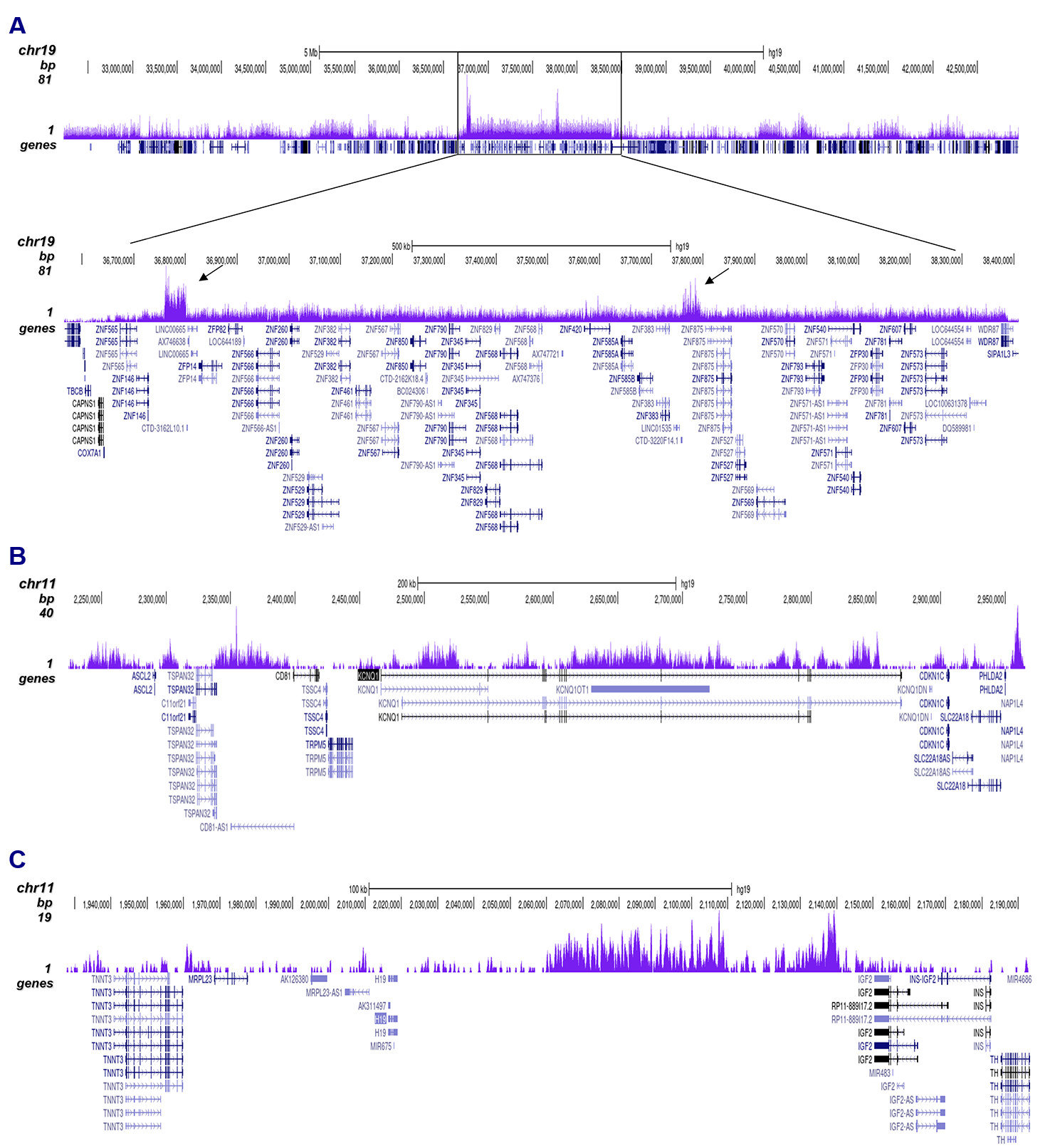
Figure 2. ChIP-seq results obtained with the antibody directed against H3K9me3
ChIP was performed with 0.5 µg of the antibody against H3K9me3 (cat. No. C15200146) on sheared chromatin from 500,000 HeLa cells using the iDeal ChIP-seq kit as described above. The IP’d DNA was subsequently analyzed on an Illumina HiSeq 2000. Library preparation, cluster generation, and sequencing were performed according to the manufacturer’s instructions. The 50 bp tags were aligned to the human genome using the BWA algorithm. Figure 2A shows the signal distribution along the long arm of chromosome 19 and a zoom-in on an enriched region containing several ZNF repeat genes. The arrows indicate two satellite repeat regions that exhibit a stronger signal. Figures 2B and 2C show the enrichment at the H19 and KCNQ1 imprinted genes.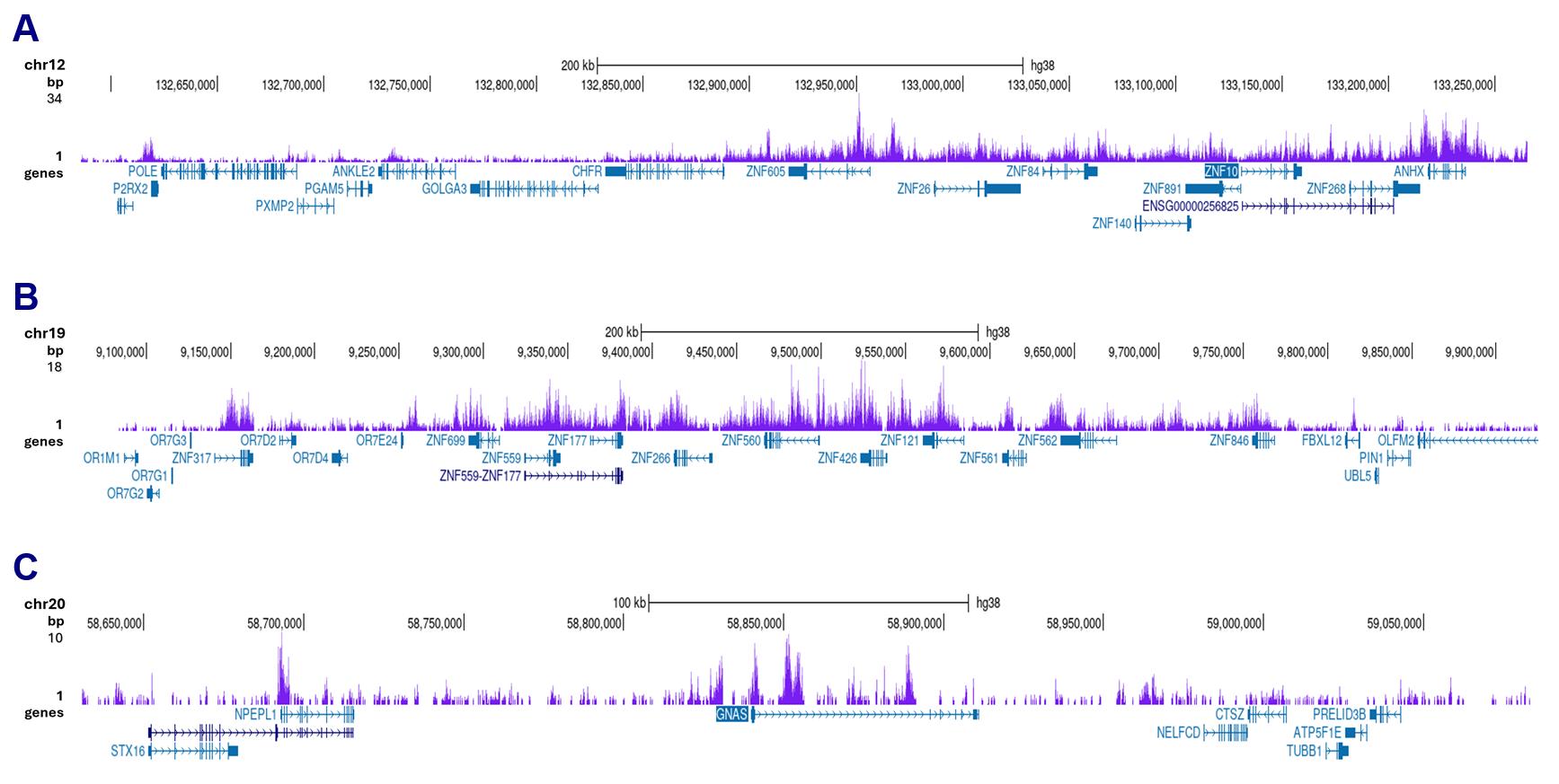
Figure 3. ut&Tag results obtained with the antibody directed against H3K9me3
Cut&Tag was performed on 50,000 K562 cells using 1 µg of the antibody against H3K9me3 (cat. No. C15200146), the pA-Tn5 transposase (C01070001), and the iDeal Cut&Tag kit (cat. No. C01070021). The libraries were subsequently analyzed on an Illumina NovaSeq sequencer (2x50 bp paired-end reads) according to the manufacturer’s instructions. The tags were aligned to the human genome (hg38) using the BWA algorithm. Figure 3 shows the peak distribution in two genomic regions on chromosomes 12 and 19 (each containing several ZNF repeat genes) and on chromosome 20 surrounding the GNAS imprinting control region (Figure 3A, B and C, respectively).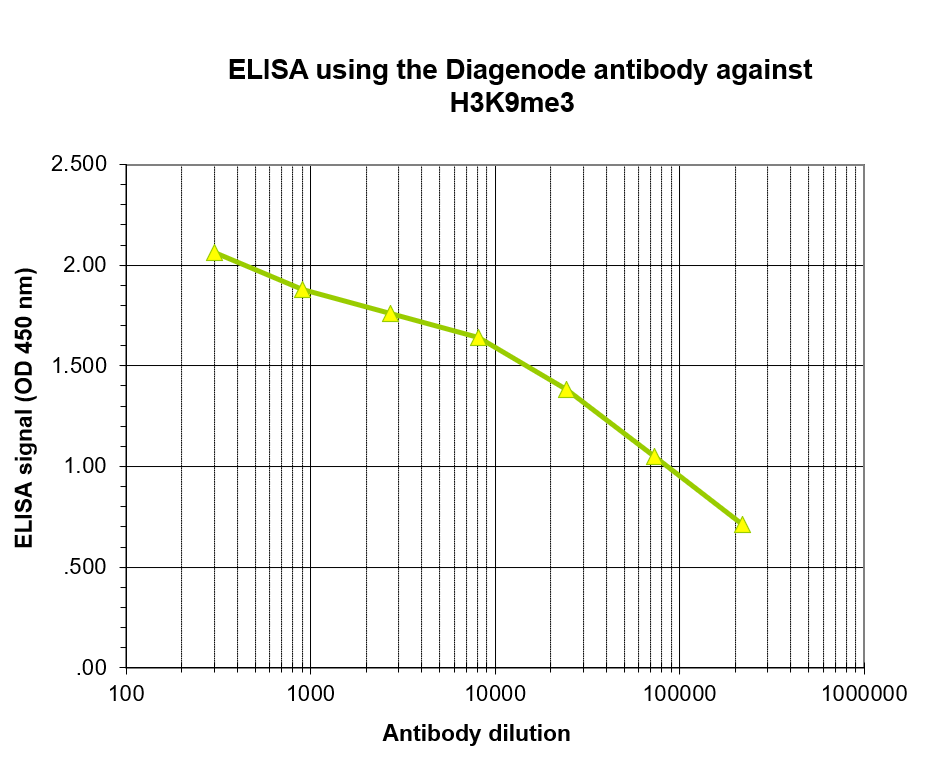
Figure 4. Determination of the antibody titer
To determine the titer of the antibody, an ELISA was performed using a serial dilution of the antibody directed against H3K9me3 (cat. No. C15200146) in antigen-coated wells. The antigen used was a peptide containing the histone modification of interest. By plotting the absorbance against the antibody dilution (Figure 4), the titer of the antibody was estimated to be 1:14,500.
Figure 5. Cross reactivity test using the monoclonal antibody directed against H3K9me3
A Dot Blot analysis was performed to test the cross-reactivity of the monoclonal antibody against H3K9me3 (cat. No. C15200146) with peptides containing different modifications of histone H3 or H4, as well as the unmodified H3K9 sequence. One hundred to 0.2 pmol of peptide containing the respective histone modification were spotted on a membrane. The antibody was used at a dilution of 1:100,000. Figure 5 shows high specificity of the antibody for the modification of interest, with some cross-reactivity to the H3K9me2 peptide.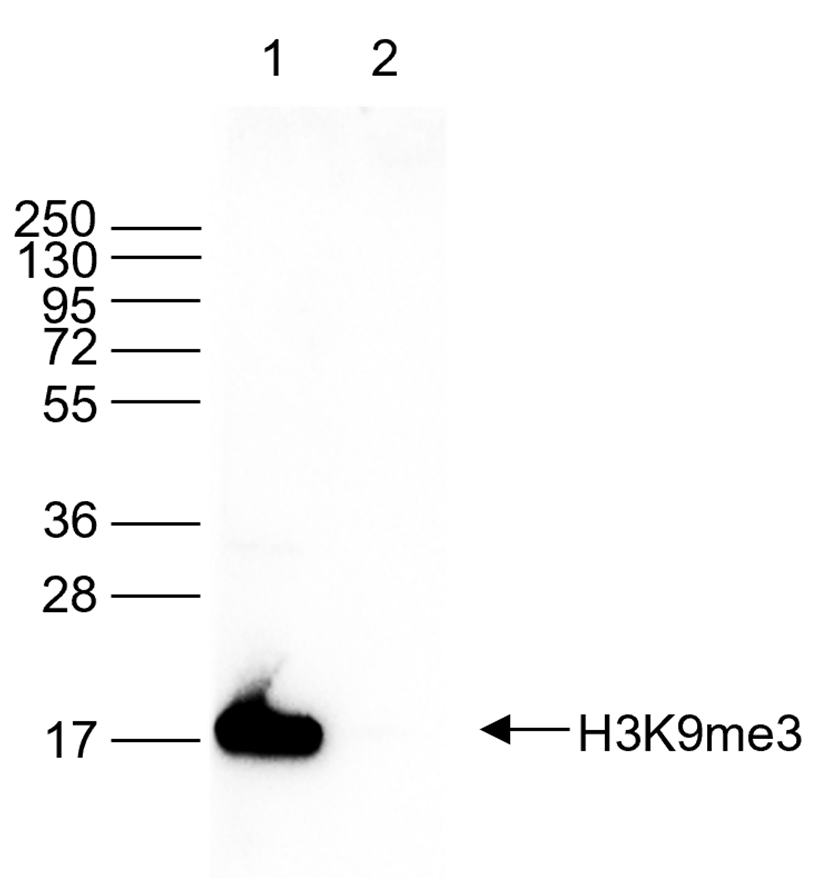
Figure 6. Western blot analysis using the monoclonal antibody directed against H3K9me3
Western blot was performed on histone extracts (15 µg, lane 1) from HeLa cells, and on 1 µg of recombinant histone H3 (lane 2) using the monoclonal antibody against H3K9me3 (cat. No. C15200146). The antibody was diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The marker (in kDa) is shown on the left; the position of the protein is indicated on the right.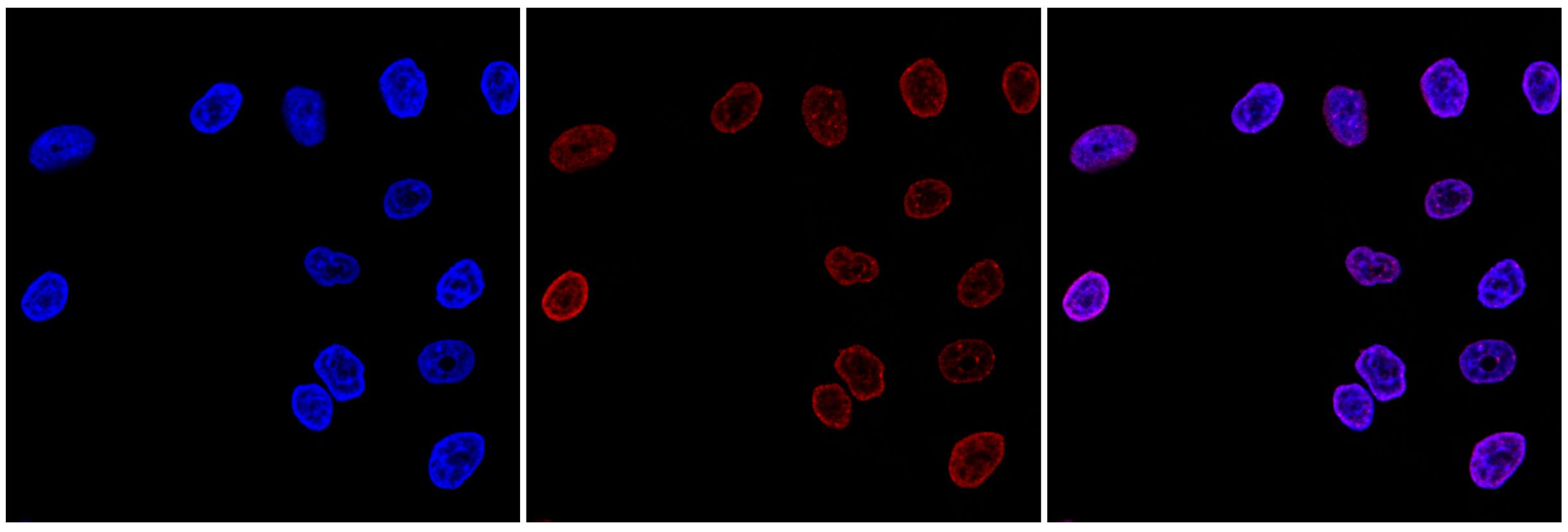
Figure 7. Immunofluorescence using the monoclonal antibody directed against H3K9me3
HeLa cells were stained with the antibody against H3K9me3 (cat. No. C15200146) and with DAPI. Cells were fixed with 4% formaldehyde for 10’ and blocked with PBS/TX-100 containing 1% BSA. The cells were immunofluorescently labeled with the H3K9me3 antibody (middle), diluted 1:500 in blocking solution, followed by an anti-mouse antibody conjugated to Alexa594. The left panel shows staining of the nuclei with DAPI. A merge of both stains is shown on the right. - Publications

How to properly cite our product/service in your work
We strongly recommend using this: H3K9me3 Antibody (sample size) (Hologic Diagenode Cat# C15200146-10 Lot# 003). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
High resolution methylation analysis of the HoxA5 regulatory region in different somatic tissues of laboratory mouse during development
Sinha P. et al.
Homeobox genes encode a group of DNA binding regulatory proteins whose key function occurs in the spatial-temporal organization of genome during embryonic development and differentiation. The role of these Hox genes during ontogenesis makes it an important model for research. HoxA5 is a member of Hox gene family pla...Methylation of the Sox9 and Oct4 promoters and its correlation with gene expression during testicular development in the laboratory mouse
Pamnani M et al.
Sox9 and Oct4 are two important regulatory factors involved in mammalian development. Sox9, a member of the group E Sox transcription factor family, has a crucial role in the development of the genitourinary system, while Oct4, commonly known as octamer binding transcription factor 4, belongs to class V of the trans...Heat shock represses rRNA synthesis by inactivation of TIF-IA and lncRNA-dependent changes in nucleosome positioning
Zhao Z et al.
Attenuation of ribosome biogenesis in suboptimal growth environments is crucial for cellular homeostasis and genetic integrity. Here, we show that shutdown of rRNA synthesis in response to elevated temperature is brought about by mechanisms that target both the RNA polymerase I (Pol I) transcription machinery and th...The histone demethylase JMJD2A/KDM4A links ribosomal RNA transcription to nutrients and growth factors availability
Salifou K, Ray S, Verrier L, Aguirrebengoa M, Trouche D, Panov KI, Vandromme M
The interplay between methylation and demethylation of histone lysine residues is an essential component of gene expression regulation and there is considerable interest in elucidating the roles of proteins involved. Here we report that histone demethylase KDM4A/JMJD2A, which is involved in the regulation of cell pr...A high-resolution imaging approach to investigate chromatin architecture in complex tissues
Linhoff MW, Garg SK, Mandel G
We present ChromATin, a quantitative high-resolution imaging approach for investigating chromatin organization in complex tissues. This method combines analysis of epigenetic modifications by immunostaining, localization of specific DNA sequences by FISH, and high-resolution segregation of nuclear compartments u...Deciphering the cryptic genome: genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites.
Wiemann P, Sieber CM, von Bargen KW, Studt L, Niehaus EM, Espino JJ, Huß K, Michielse CB, Albermann S, Wagner D, Bergner SV, Connolly LR, Fischer A, Reuter G, Kleigrewe K, Bald T, Wingfield BD, Ophir R, Freeman S, Hippler M, Smith KM, Brown DW, Proctor RH
The fungus Fusarium fujikuroi causes "bakanae" disease of rice due to its ability to produce gibberellins (GAs), but it is also known for producing harmful mycotoxins. However, the genetic capacity for the whole arsenal of natural compounds and their role in the fungus' interaction with rice remained unknown. Here, ...A novel mechanism for CTCF in the epigenetic regulation of Bax in breast cancer cells.
Méndez-Catalá CF, Gretton S, Vostrov A, Pugacheva E, Farrar D, Ito Y, Docquier F, Kita GX, Murrell A, Lobanenkov V, Klenova E.
We previously reported the association of elevated levels of the multifunctional transcription factor, CCCTC binding factor (CTCF), in breast cancer cells with the specific anti-apoptotic function of CTCF. To understand the molecular mechanisms of this phenomenon, we investigated regulation of the human Bax gene by ...