Histones are the main constituents of the protein part of chromosomes of eukaryotic cells. They are rich in the amino acids arginine and lysine and have been greatly conserved during evolution. Histones pack the DNA into tight masses of chromatin. Two core histones of each class H2A, H2B, H3 and H4 assemble and are wrapped by 146 base pairs of DNA to form one octameric nucleosome. Histone tails undergo numerous post-translational modifications, which either directly or indirectly alter chromatin structure to facilitate transcriptional activation or repression or other nuclear processes. In addition to the genetic code, combinations of the different histone modifications reveal the so-called “histone code”. Histone methylation and demethylation is dynamically regulated by respectively histone methyl transferases and histone demethylases. Trimethylation of histone H3K9 is associated with imprinted regions, satellite repeats and ZNF repeat genes.
H3K9me3 recombinant antibody and negative control (sample size)

Diagenode is pleased to offer the first recombinant antibody against H3K9me3. It has has been selected using tailored phage-display libraries, and produced in E.coli eliminating animals from the antibody-production process. This recombinant antibody shows superior specificity and affinity as well as no lot-to-lot variation.
| Lot | 001 |
|---|---|
| Concentration | 0.25 µg/µl |
| Species reactivity | Human, mouse, drosophila, yeast |
| Type | Monoclonal, Recombinant |
| Purity | affinity purified |
| Host | Fab format with human framework |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution/amount | References |
|---|---|---|
| ChIP/ChIP-seq * | 0.2-1.8 μg per ChIP | Fig 1, 2 |
| Immunofluorescence | 1:500 | Fig 3 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 0.5-5 μg per IP.
- Validation Data
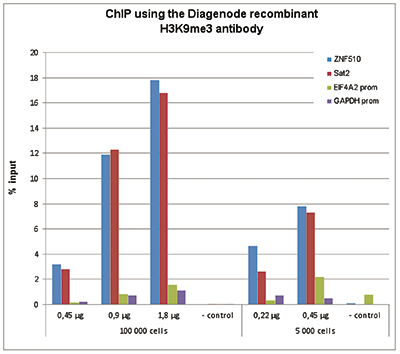
Figure 1. ChIP results obtained with the Diagenode recombinant antibody directed against H3K9me3
ChIP assays were performed using human HeLa cells, the Diagenode recombinant antibody against H3K9me3 and optimized PCR primer sets for qPCR. ChIP was performed on sheared chromatin from 100,000 and 5,000 cells with the “True MicroChIP kit (cat. No. C01010130). See page 4: Protocol for binding the recombinant H3K9me3 antibody to streptavidin- coated beads (Hattori T. et al., 2013). Different amounts of the antibody were analysed. A negative control recombinant antibody 1 or 5 μg/IP) was used as negative IP control. QPCR was performed with primers for the heterochromatin marker Sat2 and for the ZNF510 gene, used as positive controls, and for the promoters of the active EIF4A2 and GAPDH genes, used as negative controls. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).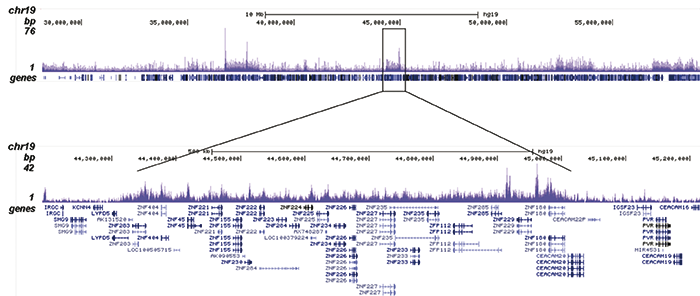
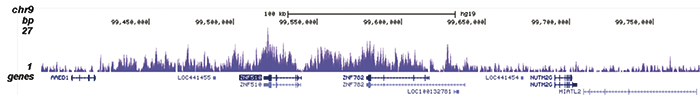
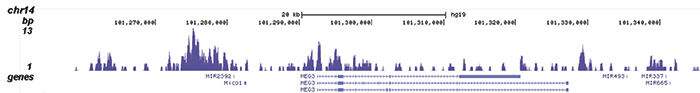
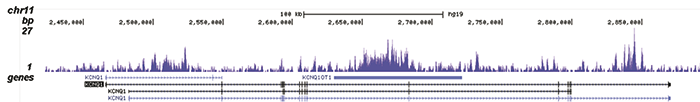
Figure 2. ChIP-seq results obtained with the Diagenode recombinant antibody directed against H3K9me3
ChIP was performed with 1.3 μg of the Diagenode antibody against H3K9me3 on sheared chromatin from 4 million K562 cells. The IP’d DNA was analysed on an Illumina Genome Analyzer. Library preparation, cluster generation and sequencing were performed according to the manufacturer’s instructions. The sequenced reads were aligned to human genome version 19 using the ELAND algorithm. Figure 2A shows the signal distribution along the long arm of chromosome 19 and a zoomin to an enriched region containing several ZNF repeat genes. Figure 2B shows the enrichment at ZNF510 and Figure 2 C and D show the enrichment at the MEG3 and KCNQ1 imprinted genes.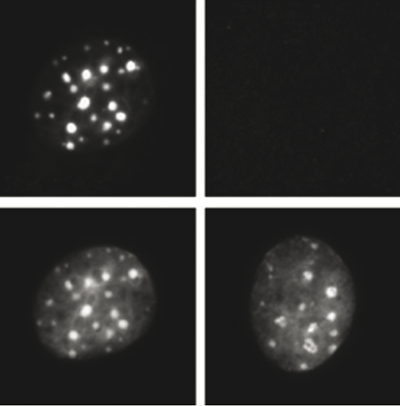
Figure 3. Immunofluorescence using the Diagenode recombinant antibody directed against H3K9me3
NIH 3T3 cells were stained with the Diagenode antibody against H3K9me3, left or with the negative control recombinant antibody, right. The bottom panel shows counterstaining of the cells with DAPI. (Hattori T. et al., 2013).Protocol for binding the recombinant H3K9me3 antibody to streptavidin-coated beads (Hattori T. et al., 2013). The recombinant H3K9me3 antibody (Cat. No. C15500003) has been validated in ChIP with the True MicroChIP kit (Cat. No. C01010130). However, since this antibody is a biotinylated Fab fragment, the protocol was slightly adapted. The protein A/G coated magnetic beads included in the True MicroChIP kit were replaced by streptavidin-coated beads to capture the recombinant antibody. The protocol below is intended for binding of the antibody to streptavidin beads for one ChIP experiment. Scale up accordingly for larger numbers of ChIP experiments.
Material required
- Dynabeads M280 Streptavidin (Invitrogen)
Alternatively Streptavidin MagneSphere paramagnetic beads (Promega) can be used - TBS containing 0.5% BSA (called TBS/BSA in the protocol)
- Biotin. Prepare a solution of 5 μM biotin in TBS containing 0.5 % BSA
- Diamag 1.5 magnetic rack (Cat No. kch-816-015)
NOTE: Please proceed with STEP 1 - Cell collection and DNA-protein crosslinking as well as STEP 2 - Cell lysis and chromatin shearing, as explained in the True MicroChIP kit protocol. In STEP 3 – Magnetic Immunoprecipitation and washes, proceed up to point 22 for Detailed protocol or point 13 for Short protocol (ie proceed up to chromatin dilution after the shearing and use this diluted chromatin at the end of the recombinant antibody binding protocol below).The protocol below is optimized for working with 100 000 cells. When using less cells, you should decrease the amount of antibody and beads to use.
- Dynabeads M280 Streptavidin (Invitrogen)
- Publications

How to properly cite our product/service in your work
We strongly recommend using this: H3K9me3 recombinant antibody and negative control (sample size) (Hologic Diagenode Cat# C15500003-10 Lot# 001). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
H3K9 dimethylation safeguards cancer cells against activation of theinterferon pathway.
Hansen A. M. et al.
Activation of interferon genes constitutes an important anticancer pathway able to restrict proliferation of cancer cells. Here, we demonstrate that the H3K9me3 histone methyltransferase (HMT) suppressor of variegation 3-9 homolog 1 (SUV39H1) is required for the proliferation of acute myeloid leukemia (AML) and find...SUV39 SET domains mediate crosstalk of heterochromatic histone marks.
Stirpe, Alessandro and Guidotti, Nora and Northall, Sarah Jand Kilic, Sinan and Hainard, Alexandre and Vadas, Oscar andFierz, Beat and Schalch, Thomas
The SUV39 class of methyltransferase enzymes deposits histone H3 lysine 9 di- and trimethylation (H3K9me2/3), the hallmark of constitutive heterochromatin. How these enzymes are regulated to mark specific genomic regions as heterochromatic is poorly understood. Clr4 is the sole H3K9me2/3 methyltransferase in the fis...A conserved RNA degradation complex required for spreading and epigenetic inheritance of heterochromatin.
Shipkovenska G, Durango A, Kalocsay M, Gygi SP, Moazed D
Heterochromatic domains containing histone H3 lysine 9 methylation (H3K9me) can be epigenetically inherited independently of underlying DNA sequence. To gain insight into the mechanisms that mediate epigenetic inheritance, we used a inducible heterochromatin formation system to perform a genetic screen for mutations...Genomic Profiling by ALaP-Seq Reveals Transcriptional Regulation by PML Bodies through DNMT3A Exclusion.
Kurihara M, Kato K, Sanbo C, Shigenobu S, Ohkawa Y, Fuchigami T, Miyanari Y
The promyelocytic leukemia (PML) body is a phase-separated nuclear structure physically associated with chromatin, implying its crucial roles in genome functions. However, its role in transcriptional regulation is largely unknown. We developed APEX-mediated chromatin labeling and purification (ALaP) to identify the ...Abo1 is required for the H3K9me2 to H3K9me3 transition in heterochromatin.
Dong W, Oya E, Zahedi Y, Prasad P, Svensson JP, Lennartsson A, Ekwall K, Durand-Dubief M
Heterochromatin regulation is critical for genomic stability. Different H3K9 methylation states have been discovered, with distinct roles in heterochromatin formation and silencing. However, how the transition from H3K9me2 to H3K9me3 is controlled is still unclear. Here, we investigate the role of the conserved brom...Native Chromatin Proteomics Reveals a Role for Specific Nucleoporins in Heterochromatin Organization and Maintenance.
Iglesias N, Paulo JA, Tatarakis A, Wang X, Edwards AL, Bhanu NV, Garcia BA, Haas W, Gygi SP, Moazed D
Spatially and functionally distinct domains of heterochromatin and euchromatin play important roles in the maintenance of chromosome stability and regulation of gene expression, but a comprehensive knowledge of their composition is lacking. Here, we develop a strategy for the isolation of native Schizosaccharom...Replication timing and epigenome remodelling are associated with the nature of chromosomal rearrangements in cancer.
Du Q, Bert SA, Armstrong NJ, Caldon CE, Song JZ, Nair SS, Gould CM, Luu PL, Peters T, Khoury A, Qu W, Zotenko E, Stirzaker C, Clark SJ
DNA replication timing is known to facilitate the establishment of the epigenome, however, the intimate connection between replication timing and changes to the genome and epigenome in cancer remain largely uncharacterised. Here, we perform Repli-Seq and integrated epigenome analyses and demonstrate that genomic reg...Automethylation-induced conformational switch in Clr4 (Suv39h) maintains epigenetic stability.
Iglesias N, Currie MA, Jih G, Paulo JA, Siuti N, Kalocsay M, Gygi SP, Moazed D
Histone H3 lysine 9 methylation (H3K9me) mediates heterochromatic gene silencing and is important for genome stability and the regulation of gene expression. The establishment and epigenetic maintenance of heterochromatin involve the recruitment of H3K9 methyltransferases to specific sites on DNA, followed by the re...Epigenetic inheritance mediated by coupling of RNAi and histone H3K9 methylation.
Yu R, Wang X, Moazed D
Histone post-translational modifications (PTMs) are associated with epigenetic states that form the basis for cell-type-specific gene expression. Once established, histone PTMs can be maintained by positive feedback involving enzymes that recognize a pre-existing histone modification and catalyse the same modificati...Unique roles for histone H3K9me states in RNAi and heritable silencing of transcription
Jih G. et al.
Heterochromatic DNA domains have important roles in the regulation of gene expression and maintenance of genome stability by silencing repetitive DNA elements and transposons. From fission yeast to mammals, heterochromatin assembly at DNA repeats involves the activity of small noncoding RNAs (sRNAs) associated with ...