| WB Western blot : The quality of antibodies used in this technique is crucial for correct and specific protein identification. Diagenode offers huge selection of highly sensitive and specific western blot-validated antibodies. Learn more about: Load... Read more |
| ELISA Enzyme-linked immunosorbent assay. Read more |
| IF Immunofluorescence: Diagenode offers huge selection of highly sensitive antibodies validated in IF. Immunofluorescence using the Diagenode monoclonal antibody directed against CRISPR/Cas9 HeLa cells transfected with a Cas9 expression vector (... Read more |
| ChIP-qPCR (ab) Read more |
H3K9me2 monoclonal antibody (sample size)

Monoclonal antibody raised in mouse against histone H3, dimethylated at lysine 9 (H3K9me2), using a KLH-conjugated synthetic peptide.
| Lot | 001-013 |
|---|---|
| Concentration | 1.0 µg/µl |
| Species reactivity | Human, zebrafish, Arabidopsis, silena latifolia: positive. Other species: not tested. |
| Type | Monoclonal |
| Purity | Protein A purified monoclonal antibody. |
| Host | Mouse |
| Storage Conditions | Store at -20°C; for long storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Storage Buffer | PBS containing 0.05% azide and 0.05% ProClin 300. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP * | 3 µg/ChIP | Fig 1 |
| ELISA | 1:1,000 | Fig 2 |
| Western Blotting | 1:1,000 | Fig 3 |
| Immunofluorescence | 1:500 | Fig 4 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-5 µg per IP.
- Validation data
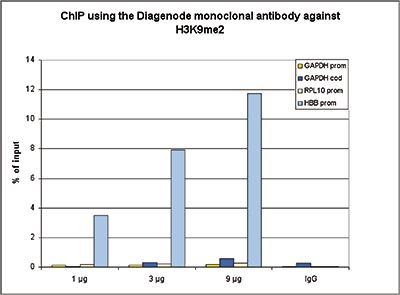
Figure 1. ChIP results obtained with the Diagenode monoclonal antibody directed against H3K9me2
ChIP assays were performed using HeLa cells, the monoclonal antibody against H3K9me2 (Cat. No. C15200154) and optimized PCR primer sets for qPCR. Chromatin was sheared with the Diagenode Bioruptor using the “Shearing ChIP” kit (Cat. No. C01020021). ChIP was performed with the “OneDay ChIP” kit (Cat. No. C01010080), using sheared chromatin from 1.6 million cells. A titration of the antibody consisting of 1, 3 and 9 µg per ChIP experiment was analysed. IgG (5 µg/IP) was used as negative IP control. QPCR was performed with primers for the promoter and the coding region of the GAPDH gene, and for the RPL10 and HBB promoters. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).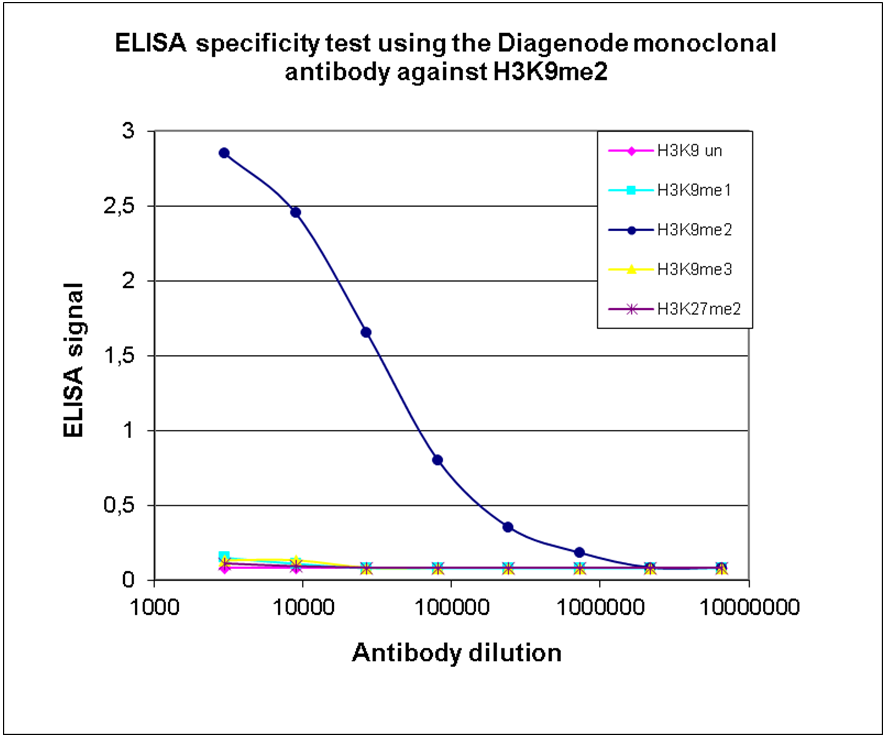
Figure 2. Cross reactivity of the Diagenode monoclonal antibody directed against H3K9me2
To test the specificity an ELISA was performed using a serial dilution of the Diagenode monoclonal antibody against H3K9me2 (cat. No. C15200154). The wells were coated with peptides containing the unmodified H3K9 as well as the mono-, di- and trimethylated H3K9 and the dimethylated H3K27. Figure 2 shows a high specificity of the antibody for the modification of interest.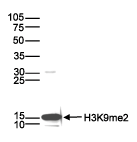
Figure 3. Western blot analysis using the Diagenode monoclonal antibody directed against H3K9me2
Histone extracts (15 µg) from HeLa cells were analysed by Western blot using the Diagenode monoclonal antibody against H3K9me2 (Cat. No. C15200154) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left.
Figure 4. Immunofluorescence using the Diagenode monoclonal antibody directed against H3K9me2
HeLa cells were stained with the Diagenode antibody against H3K9me2 (Cat. No. C15200154) and with DAPI. Cells were fixed with 4% formaldehyde for 10’ and blocked with PBS/TX-100 containing 5% normal goat serum and 1% BSA. The cells were immunofluorescently labelled with the H3K9me2 antibody (left) diluted 1:500 in blocking solution followed by an anti-mouse antibody conjugated to Alexa594. The middle panel shows staining of the nuclei with DAPI. A merge of the two stainings is shown on the right. - 出版物

How to properly cite our product/service in your work
We strongly recommend using this: H3K9me2 monoclonal antibody (sample size) (Hologic Diagenode Cat# C15200154-10 Lot# 001-013). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
Histone lysine demethylase inhibition reprograms prostate cancermetabolism and mechanics.
Chianese Ugo and Papulino Chiara and Passaro Eugenia andEvers Tom Mj and Babaei Mehrad and Toraldo Antonella andDe Marchi Tommaso and Niméus Emma and Carafa Vincenzo andNicoletti Maria Maddalena and Del Gaudio Nunzio andIaccarino Nunzia an
OBJECTIVE: Aberrant activity of androgen receptor (AR) is the primary cause underlying development and progression of prostate cancer (PCa) and castration-resistant PCa (CRPC). Androgen signaling regulates gene transcription and lipid metabolism, facilitating tumor growth and therapy resistance in early and advanced...Histone post-translational modifications in Silene latifolia X and Y chromosomes suggest a mammal-like dosage compensation system
Luis Rodríguez Lorenzo José, Hubinský Marcel, Vyskot Boris, Hobza Roman
Silene latifolia is a model organism to study evolutionary young heteromorphic sex chromosome evolution in plants. Previous research indicates a Y-allele gene degeneration and a dosage compensation system already operating. Here, we propose an epigenetic approach based on analysis of several histone post-translation...The histone methyltransferase DOT1L is required for proper DNA damage response, DNA repair, and modulates chemotherapy responsiveness.
Kari V, Raul SK, Henck JM, Kitz J, Kramer F, Kosinsky RL, Übelmesser N, Mansour WY, Eggert J, Spitzner M, Najafova Z, Bastians H, Grade M, Gaedcke J, Wegwitz F, Johnsen SA
BACKGROUND: Disruptor of telomeric silencing 1-like (DOT1L) is a non-SET domain containing methyltransferase known to catalyze mono-, di-, and tri-methylation of histone 3 on lysine 79 (H3K79me). DOT1L-mediated H3K79me has been implicated in chromatin-associated functions including gene transcription, heterochromati...Role of Annexin gene and its regulation during zebrafish caudal fin regeneration
Saxena S, Purushothaman S, Meghah V, Bhatti B, Poruri A, Meena Lakshmi MG, Sarath Babu N, Murthy CL, Mandal KK, Kumar A, Idris MM
The molecular mechanism of epimorphic regeneration is elusive due to its complexity and limitation in mammals. Epigenetic regulatory mechanisms play a crucial role in development and regeneration. This investigation attempted to reveal the role of epigenetic regulatory mechanisms, such as histone H3 and H4 lysine ac...Independent Mechanisms Target SMCHD1 to Trimethylated Histone H3 Lysine 9-Modified Chromatin and the Inactive X Chromosome
Brideau NJ, Coker H, Gendrel AV, Siebert CA, Bezstarosti K, Demmers J, Poot RA, Nesterova TB, Brockdorff N
The chromosomal protein SMCHD1 plays an important role in epigenetic silencing at diverse loci, including the inactive X chromosome, imprinted genes, and the facioscapulohumeral muscular dystrophy locus. Although homology with canonical SMC family proteins suggests a role in chromosome organization, the mechanisms u...A gene expression signature identifying transient DNMT1 depletion as a causal factor of cancer-germline gene activation in melanoma
Charles De Smet
Many human tumors show aberrant activation of a group of germline-specific genes, termed cancer-germline (CG) genes, several of which appear to exert oncogenic functions. Although activation of CG genes in tumors has been linked to promoter DNA demethylation, the mechanisms underlying this epigenetic alteration rema...Epigenetic hierarchy within the MAGEA1 cancer-germline gene: promoter DNA methylation dictates local histone modifications.
Cannuyer J, Loriot A, Parvizi GK, De Smet C
Gene MAGEA1 belongs to a group of human germline-specific genes that rely on DNA methylation for repression in somatic tissues. Many of these genes, termed cancer-germline (CG) genes, become demethylated and activated in a wide variety of tumors, where they encode tumor-specific antigens. The process leading to DNA ...