RNA polymerase II (pol II) is a key enzyme in the regulation and control of gene transcription. It is able to unwind the DNA double helix, synthesize RNA, and proofread the result. Pol II is a complex enzyme, consisting of 12 subunits, of which the B1 subunit (UniProt/Swiss-Prot entry P24928) is the largest. Together with the second largest subunit, B1 forms the catalytic core of the RNA polymerase II transcription machinery.
Pol II monoclonal antibody (sample size)

Alternative names: POLR2A, RPB1, POLR2, RPOL2
Monoclonal antibody raised in mouse against the YSPTSPS repeat in the B1 subunit of RNA polymerase II.
| Lot | 001-15 |
|---|---|
| Concentration | 1.0 µg/µl |
| Species reactivity | Human, Xenopus, yeast: positive. Other species: not tested. |
| Type | Monoclonal ChIP grade, ChIP-seq grade, CUT&Tag grade |
| Purity | Protein A purified monoclonal antibody. |
| Host | Mouse |
| Storage Conditions | Store at -20°C; for long storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Storage Buffer | PBS containing 0.05% azide. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 1 µg/ChIP | Fig 1, 2 |
| CUT&Tag | 1 µg | Fig 3 |
| ELISA | 1:3,000 | Fig 4 |
| Western Blotting | 1:1,000 | Fig 5, 6 |
| Immunofluorescence | 1:500 | Fig 7 |
* Please note that the optimal antibody amount per ChIP should be determined by the end-user. We recommend testing 1-5 µg per IP.
- Validation Data
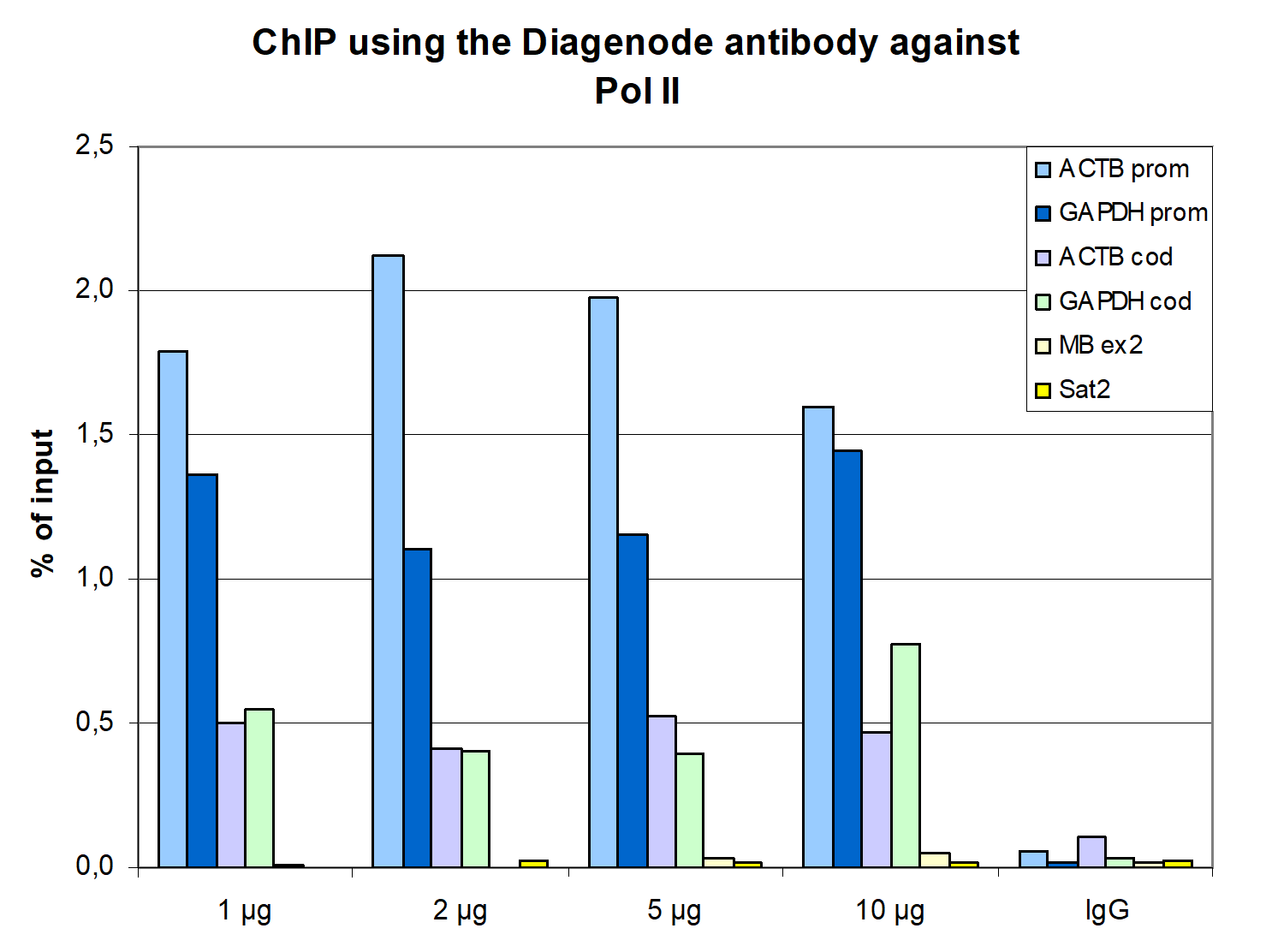
Figure 1. ChIP results obtained with the monoclonal antibody directed against Pol II
ChIP assays were performed using human HeLa cells, the monoclonal antibody against Pol II (cat. No. C15200004), and optimized PCR primer pairs for qPCR. ChIP was performed with the iDeal ChIP-seq kit (cat. No. C01010051), using sheared chromatin from 1 million cells. A titration consisting of 1, 2, 5, and 10 µg of antibody per ChIP experiment was analyzed. IgG (2 µg/IP) was used as a negative IP control. Quantitative PCR was performed with primers specific for the promoter and the coding region of the constitutively expressed GAPDH and ACTB genes, used as positive controls, and for exon 2 of the inactive myoglobin (MB) gene and the Sat2 satellite repeat, used as negative controls. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).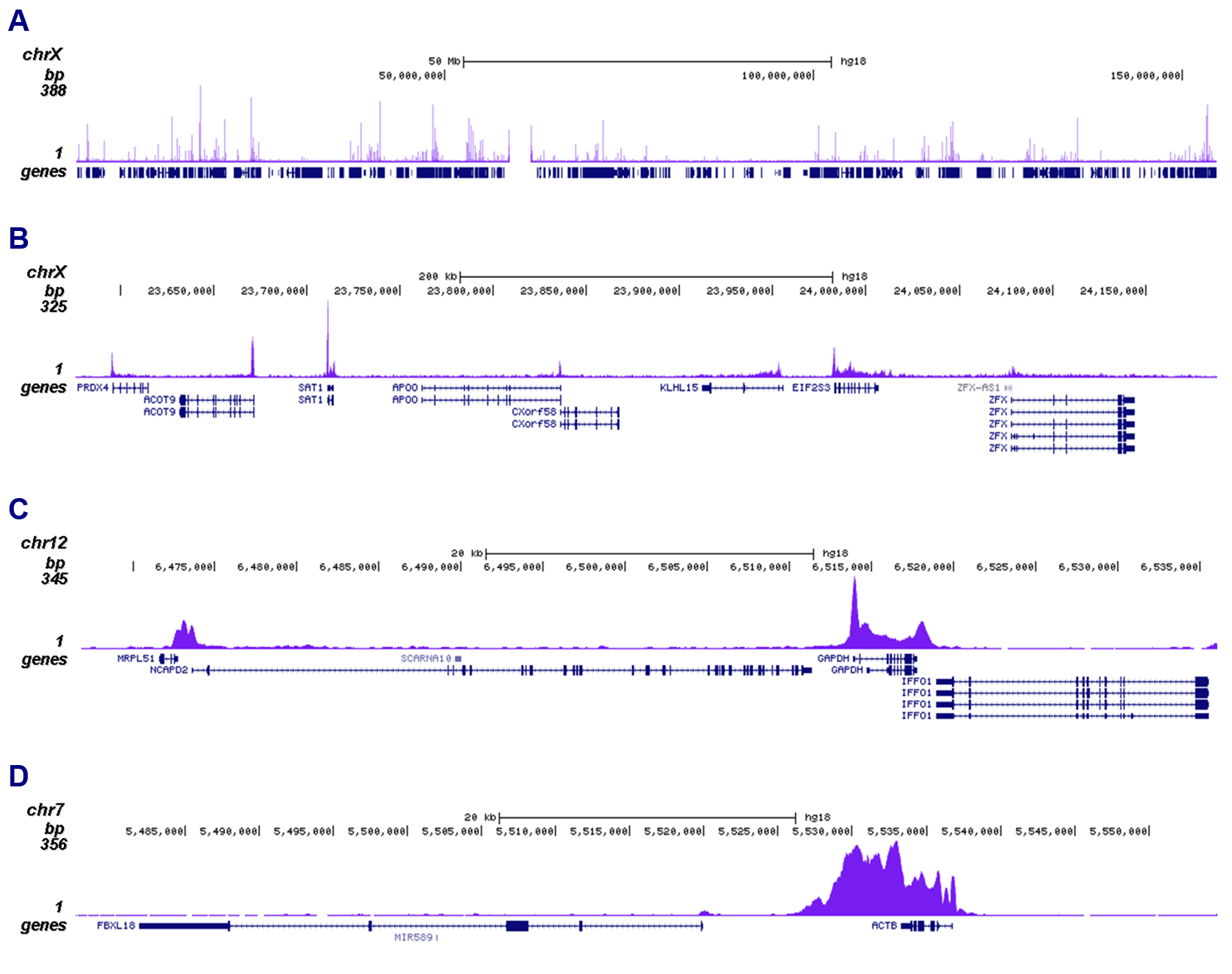
Figure 2. ChIP-seq results obtained with the monoclonal antibody directed against Pol II
ChIP was performed on sheared chromatin from 1 million HeLaS3 cells using 1 µg of the antibody against Pol II (cat. No. C15200004) as described above. The IP’d DNA was subsequently analyzed on an Illumina Genome Analyzer. Library preparation, cluster generation, and sequencing were performed according to the manufacturer’s instructions. The 36 bp tags were aligned to the human genome using the ELAND algorithm. Figure 2 shows the peak distribution across the complete sequence and a 400 kb region of the X-chromosome (Figure 2A and 2B, respectively), and in two genomic regions surrounding the GAPDH and ACTB positive control genes (Figure 2C and 2D).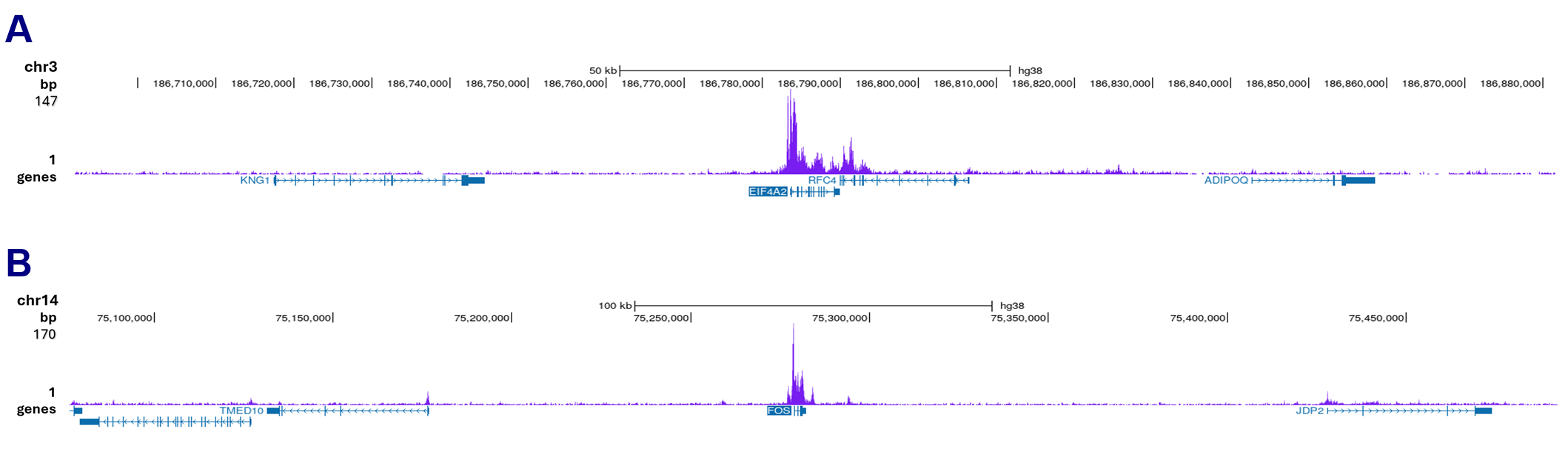
Figure 3. Cut&Tag results obtained with the monoclonal antibody directed against Pol II
Cut&Tag was performed on 50,000 K562 cells using 1 µg of the monoclonal antibody against Pol II (cat. No. C15200004), the pA-Tn5 transposase (C01070001), and the Universal Cut&Tag kit (cat. No. C01070024). The libraries were subsequently analyzed on an Illumina NovaSeq sequencer (2x50 paired-end reads) according to the manufacturer’s instructions. The tags were aligned to the human genome (hg38) using the BWA algorithm. Figure 3 shows the peak distribution in two genomic regions on chromosomes 3 and 14 (Figure 3A and 3B, respectively).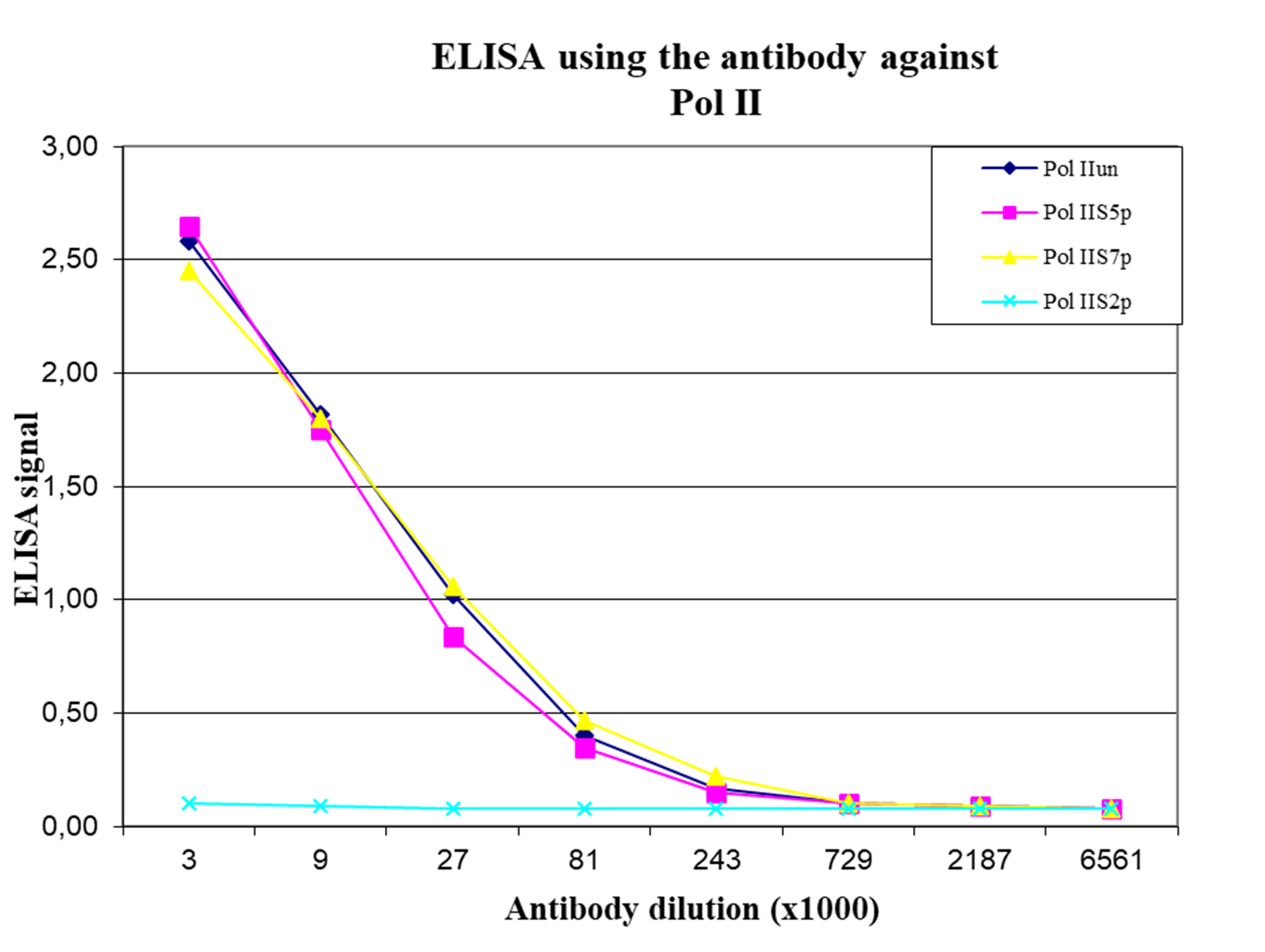
Figure 4. Cross-reactivity of the monoclonal antibody directed against Pol II
To test the specificity, an ELISA was performed using a serial dilution of the monoclonal antibody against Pol II (cat. No. C15200004). The wells were coated with peptides containing the unmodified C-terminal repeat sequence as well as different phosphorylated peptides. Figure 4 shows that the antibody recognizes the unphosphorylated Pol II as well as most phosphorylated forms.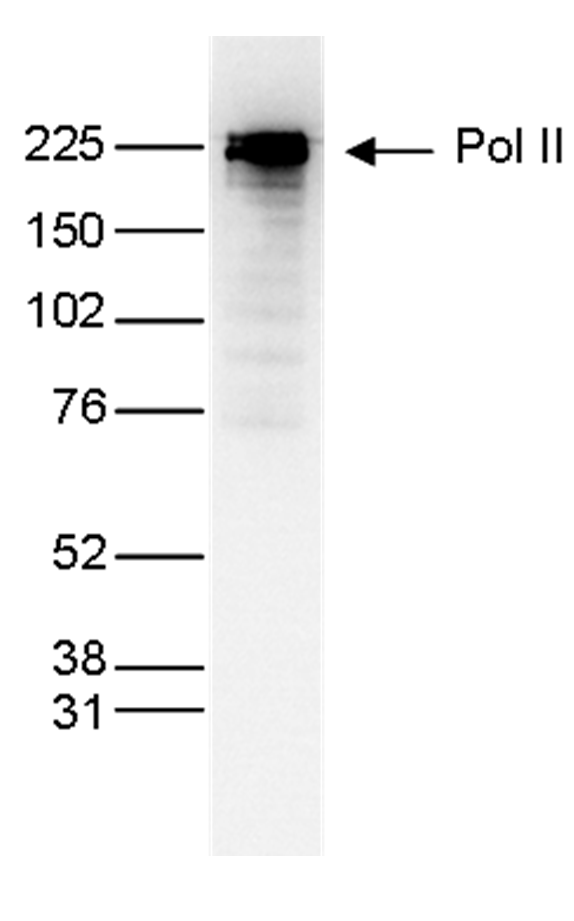
Figure 5. Western blot analysis using the monoclonal antibody directed against Pol IIantibody directed against Pol II Nuclear extracts (25 µg) from HeLa cells were analyzed by Western blot, using the monoclonal antibody against Pol II (cat. No. C15200004) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left.
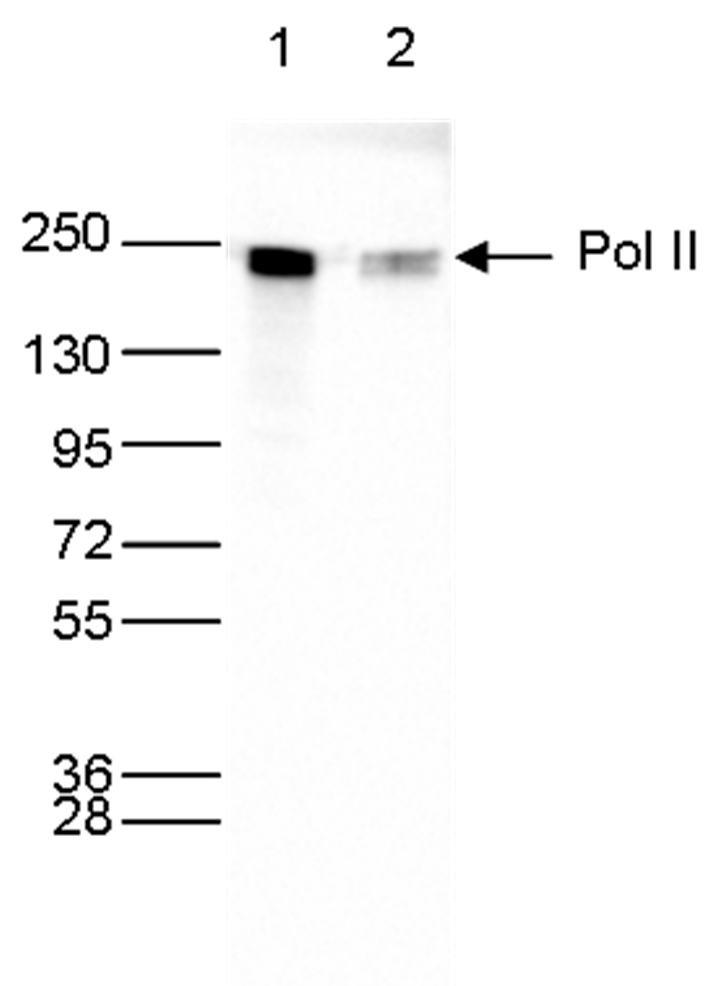
Figure 6. Western blot analysis using the monoclonal antibody directed against Pol II
Whole cell extracts (40 µg) from HeLa cells transfected with Pol II siRNA (lane 2) and from an untransfected control (lane 1) were analyzed by Western blot using the antibody against Pol II (cat. No. C15200004) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left.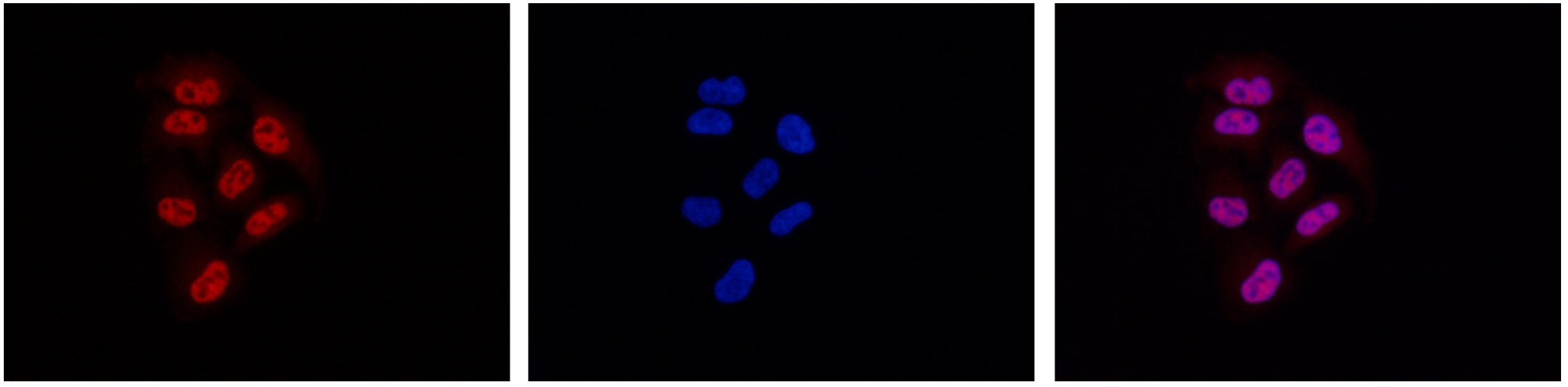
Figure 7. Immunofluorescence using the monoclonal antibody directed against Pol II
HeLa cells were stained with the antibody against Pol II (cat. No. C15200004) and with DAPI. Cells were fixed with methanol and blocked with PBS/TX-100 containing 5% normal goat serum and 1% BSA. The cells were immunofluorescently labeled with the Pol II antibody (left) diluted 1:500 in blocking solution, followed by an anti-mouse antibody conjugated to Alexa594. The middle panel shows staining of the nuclei with DAPI. A merge of the two stains is shown on the right. - 出版物

How to properly cite our product/service in your work
We strongly recommend using this: Pol II monoclonal antibody (sample size) (Hologic Diagenode Cat# C15200004-10 Lot# 001-15). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
Sox8 is essential for vertebrate gastrulation
Moreira, Sofia et al.
Gastrulation is a fundamental developmental process during which germ layers are formed and the body axes are defined by the precise orchestration of cell movements and fate specification. Here, we identify the SOXE transcription factor Sox8 as a pivotal regulator of Xenopus laevis gastrulation. We show that Sox8 ...HIRA supports hepatitis B virus minichromosome establishment andtranscriptional activity in infected hepatocytes.
Locatelli M. et al.
BACKGROUND \& AIMS: Upon Hepatitis B virus (HBV) infection, partially double stranded viral DNA converts into a covalently-closed-circular chromatinized episomal structure (cccDNA). This form represents the long-lived genomic reservoir responsible for viral persistence in the infected liver. While the involvemen...NR4A1 regulates expression of immediate early genes, suppressingreplication stress in cancer.
Guo Hongshan et al.
Deregulation of oncogenic signals in cancer triggers replication stress. Immediate early genes (IEGs) are rapidly and transiently expressed following stressful signals, contributing to an integrated response. Here, we find that the orphan nuclear receptor NR4A1 localizes across the gene body and 3' UTR of IEGs, wher...Vulnerability of drug-resistant EML4-ALK rearranged lung cancer to transcriptional inhibition.
Paliouras AR, Buzzetti M, Shi L, Donaldson IJ, Magee P, Sahoo S, Leong HS, Fassan M, Carter M, Di Leva G, Krebs MG, Blackhall F, Lovly CM, Garofalo M
A subset of lung adenocarcinomas is driven by the EML4-ALK translocation. Even though ALK inhibitors in the clinic lead to excellent initial responses, acquired resistance to these inhibitors due to on-target mutations or parallel pathway alterations is a major clinical challenge. Exploring these mechanisms of resis...High density of unrepaired genomic ribonucleotides leads to Topoisomerase 1-mediated severe growth defects in absence of ribonucleotide reductase.
Cerritelli SM, Iranzo J, Sharma S, Chabes A, Crouch RJ, Tollervey D, Hage AE
Cellular levels of ribonucleoside triphosphates (rNTPs) are much higher than those of deoxyribonucleoside triphosphates (dNTPs), thereby influencing the frequency of incorporation of ribonucleoside monophosphates (rNMPs) by DNA polymerases (Pol) into DNA. RNase H2-initiated ribonucleotide excision repair (RER) effic...Recombination may occur in the absence of transcription in the immunoglobulin heavy chain recombination centre.
Oudinet C, Braikia FZ, Dauba A, Khamlichi AA
Developing B cells undergo V(D)J recombination to generate a vast repertoire of Ig molecules. V(D)J recombination is initiated by the RAG1/RAG2 complex in recombination centres (RCs), where gene segments become accessible to the complex. Whether transcription is the causal factor of accessibility or whether it is a ...Recurrent SMARCB1 Mutations Reveal a Nucleosome Acidic Patch Interaction Site That Potentiates mSWI/SNF Complex Chromatin Remodeling.
Valencia AM, Collings CK, Dao HT, St Pierre R, Cheng YC, Huang J, Sun ZY, Seo HS, Mashtalir N, Comstock DE, Bolonduro O, Vangos NE, Yeoh ZC, Dornon MK, Hermawan C, Barrett L, Dhe-Paganon S, Woolf CJ, Muir TW, Kadoch C
Mammalian switch/sucrose non-fermentable (mSWI/SNF) complexes are multi-component machines that remodel chromatin architecture. Dissection of the subunit- and domain-specific contributions to complex activities is needed to advance mechanistic understanding. Here, we examine the molecular, structural, and genome-wid...The SS18-SSX Fusion Oncoprotein Hijacks BAF Complex Targeting and Function to Drive Synovial Sarcoma.
McBride MJ, Pulice JL, Beird HC, Ingram DR, D'Avino AR, Shern JF, Charville GW, Hornick JL, Nakayama RT, Garcia-Rivera EM, Araujo DM, Wang WL, Tsai JW, Yeagley M, Wagner AJ, Futreal PA, Khan J, Lazar AJ, Kadoch C
Synovial sarcoma (SS) is defined by the hallmark SS18-SSX fusion oncoprotein, which renders BAF complexes aberrant in two manners: gain of SSX to the SS18 subunit and concomitant loss of BAF47 subunit assembly. Here we demonstrate that SS18-SSX globally hijacks BAF complexes on chromatin to activate an SS transcript...The Polycomb-Dependent Epigenome Controls β Cell Dysfunction, Dedifferentiation, and Diabetes.
Lu TT, Heyne S, Dror E, Casas E, Leonhardt L, Boenke T, Yang CH, Sagar , Arrigoni L, Dalgaard K, Teperino R, Enders L, Selvaraj M, Ruf M, Raja SJ, Xie H, Boenisch U, Orkin SH, Lynn FC, Hoffman BG, Grün D, Vavouri T, Lempradl AM, Pospisilik JA
To date, it remains largely unclear to what extent chromatin machinery contributes to the susceptibility and progression of complex diseases. Here, we combine deep epigenome mapping with single-cell transcriptomics to mine for evidence of chromatin dysregulation in type 2 diabetes. We find two chromatin-state signat...Epigenetic regulation of vascular NADPH oxidase expression and reactive oxygen species production by histone deacetylase-dependent mechanisms in experimental diabetes.
Manea SA, Antonescu ML, Fenyo IM, Raicu M, Simionescu M, Manea A
Reactive oxygen species (ROS) generated by up-regulated NADPH oxidase (Nox) contribute to structural-functional alterations of the vascular wall in diabetes. Epigenetic mechanisms, such as histone acetylation, emerged as important regulators of gene expression in cardiovascular disorders. Since their role in diabete...CDK8/19 Mediator kinases potentiate induction of transcription by NFκB
Chen M. et al.
The nuclear factor-κB (NFκB) family of transcription factors has been implicated in inflammatory disorders, viral infections, and cancer. Most of the drugs that inhibit NFκB show significant side effects, possibly due to sustained NFκB suppression. Drugs affecting induced, but not basal, NF&k...Functional incompatibility between the generic NF-κB motif and a subtype-specific Sp1III element drives the formation of HIV-1 subtype C viral promoter
Verma A et al.
Of the various genetic subtypes of HIV-1, HIV-2 and SIV, only in subtype C of HIV-1, a genetically variant NF-κB binding site is found at the core of the viral promoter in association with a subtype-specific Sp1III motif. How the subtype-associated variations in the core transcription factor binding sites (TFB...Embryonic transcription is controlled by maternally defined chromatin state
Hontelez S et al.
Histone-modifying enzymes are required for cell identity and lineage commitment, however little is known about the regulatory origins of the epigenome during embryonic development. Here we generate a comprehensive set of epigenome reference maps, which we use to determine the extent to which maternal factors shape c...iRNA-seq: computational method for genome-wide assessment of acute transcriptional regulation from total RNA-seq data.
Madsen JG, Schmidt SF, Larsen BD, Loft A, Nielsen R, Mandrup S
RNA-seq is a sensitive and accurate technique to compare steady-state levels of RNA between different cellular states. However, as it does not provide an account of transcriptional activity per se, other technologies are needed to more precisely determine acute transcriptional responses. Here, we have developed an e...PRAME induces genomic instability in uveal melanoma
Harbour J. W. et al.
PRAME is a CUL2 ubiquitin ligase subunit that is normally expressed in the testis but becomes aberrantly overexpressed in many cancer types in association with aneuploidy and metastasis. Here, we show that PRAME is expressed predominantly in spermatogonia around the time of meiotic crossing-over in coordination with... - 関連商品