Histones are the main constituents of the protein part of chromosomes of eukaryotic cells. They are rich in the amino acids arginine and lysine and have been greatly conserved during evolution. Histones pack the DNA into tight masses of chromatin. Two core histones of each class H2A, H2B, H3, and H4 assemble and are wrapped by 146 base pairs of DNA to form an octameric nucleosome. Histone tails undergo numerous posttranslational modifications, which either directly or indirectly alter chromatin structure to facilitate transcriptional activation or repression or other nuclear processes. In addition to the genetic code, combinations of the different histone modifications reveal the so-called “histone code.” Histone methylation and demethylation are dynamically regulated by histone methyltransferases and histone demethylases, respectively.
H3K4me3 Antibody

Monoclonal antibody raised in mouse against histone H3, trimethylated at lysine 4 (H3K4me3), using a KLH-conjugated synthetic peptide.
| Lot | 001-19 |
|---|---|
| Concentration | 1.0 µg/µl |
| Species reactivity | Human, Mouse, Nematodes, Arabidopsis: positive. Other species: not tested. |
| Type | Monoclonal, ChIP-grade, ChIP-seq grade, CUT&Tag grade |
| Purity | Protein A purified monoclonal antibody. |
| Host | Mouse |
| Storage Conditions | Store at -20°C. For long-term storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Storage Buffer | PBS containing 0.05% azide. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 2 μg/ChIP | Fig 1,2 |
| CUT&TAG | 1 μg | Fig 3 |
| ELISA | 1:3,000 | Fig 4 |
| Western Blotting | 1:1,000 | Fig 5 |
| Immunofluorescence | 1:500 | Fig 6 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-5 μg per IP.
- Validation Data
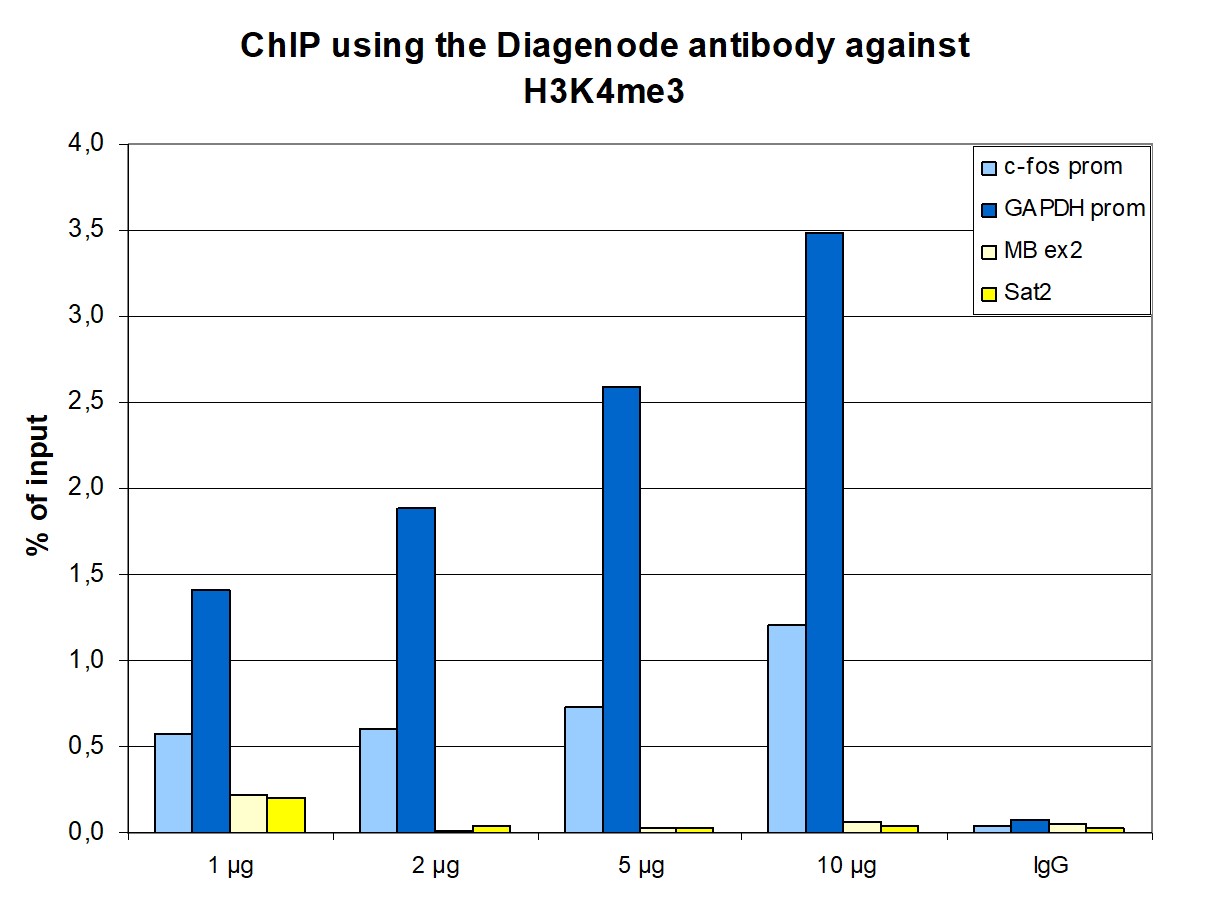
Figure 1. ChIP results obtained with the monoclonal antibody directed against H3K4me3
ChIP assays were performed using human HeLa cells, the monoclonal antibody against H3K4me3 (cat. no. C15200152), and optimized PCR primer pairs for qPCR. ChIP was performed with the iDeal ChIP-seq kit (cat. no. C01010051), using sheared chromatin from one million cells. A titration consisting of 1, 2, 5, and 10 μg of antibody per ChIP experiment was analyzed. IgG (2 μg/IP) was used as a negative IP control. Quantitative PCR was performed with primers specific for the promoter of the constitutively expressed GAPDH and c-fos genes, used as positive controls, and for exon 2 of the inactive myoglobin (MB) gene and the Sat2 satellite repeat, used as negative controls. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis). These results are in accordance with the observation that trimethylation of K4 at histone H3 is associated with the promoters of active genes.
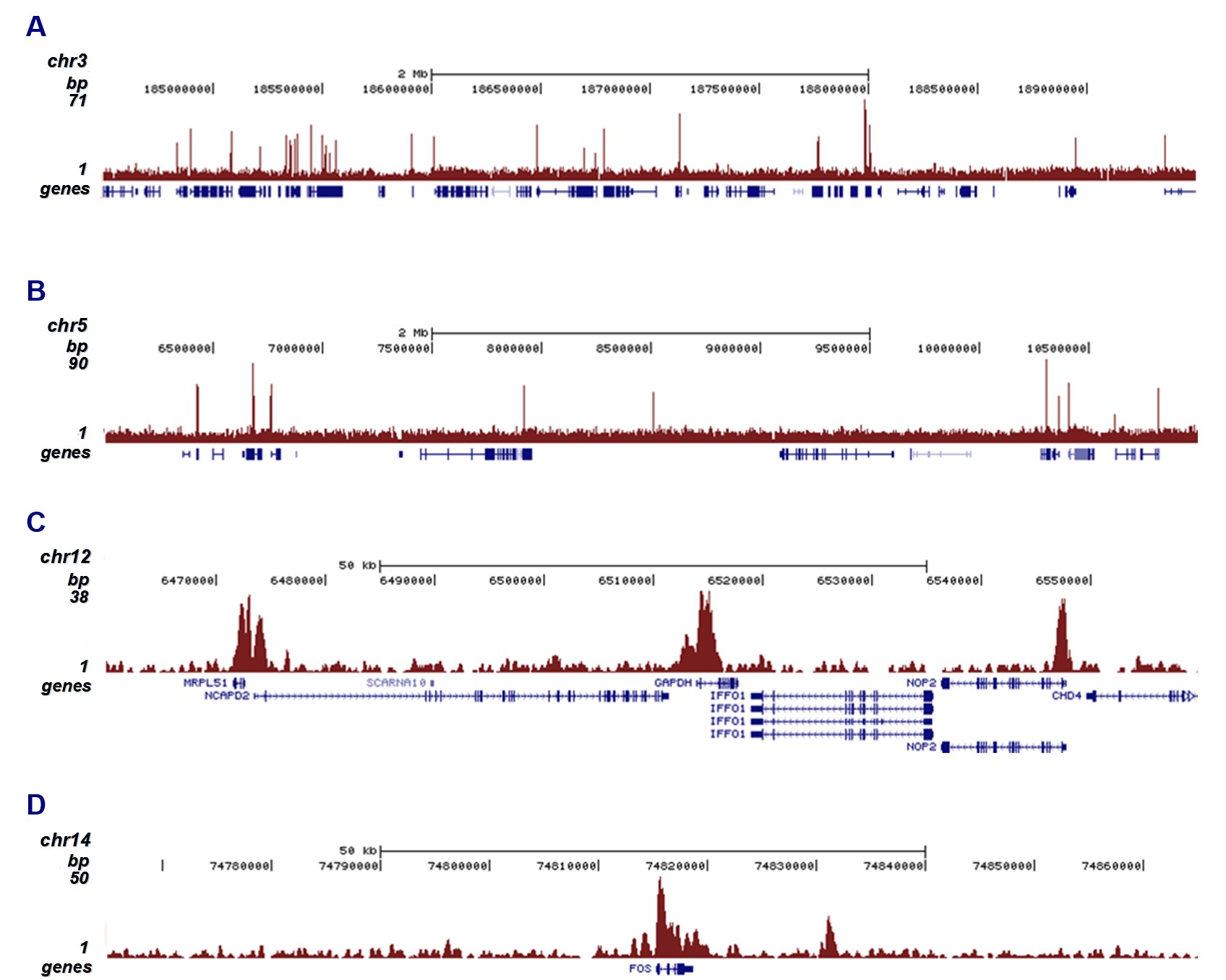
Figure 2. ChIP-seq results obtained with the monoclonal antibody directed against H3K4me3
ChIP was performed on sheared chromatin from one million HeLaS3 cells using 2 μg of the antibody against H3K4me3 (cat. no. C15200152) as described above. The IP'd DNA was subsequently analyzed on an Illumina Genome Analyzer. Library preparation, cluster generation, and sequencing were performed according to the manufacturer's instructions. The 36 bp tags were aligned to the human genome using the ELAND algorithm. Figure 2 shows the peak distribution along two 5 Mb regions of chromosomes 3 and 5 (Figure 2A and 2B, respectively) and in two 100 kb regions surrounding the GAPDH and c-fos positive control genes (Figure 2C and 2D). These results clearly show an enrichment of H3K4me3 at the promoters of active genes.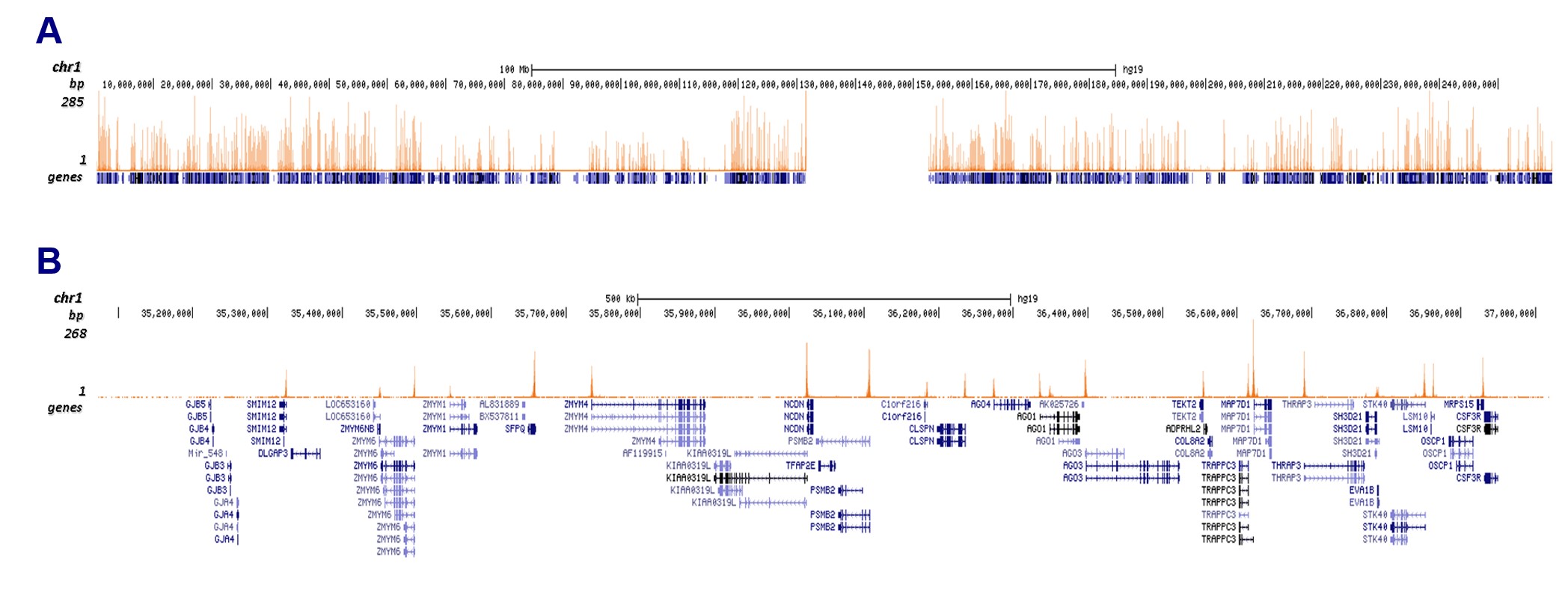
Figure 3. CUT&Tag results obtained with the monoclonal antibody directed against H3K4me3
CUT&Tag was performed on 50,000 K562 cells using 1 μg of the monoclonal antibody against H3K4me3 (cat. no. C15200152) and the iDeal CUT&Tag kit (C01070020). The libraries were subsequently analyzed on an Illumina NextSeq 500 sequencer (2 × 75 paired-end reads) according to the manufacturer's instructions. The tags were aligned to the human genome (hg19) using the BWA algorithm. Figure 3 shows the peak distribution along the complete sequence and a 1.5 Mb zoom-in of chromosome 1 (Figure 3A and 3B, respectively).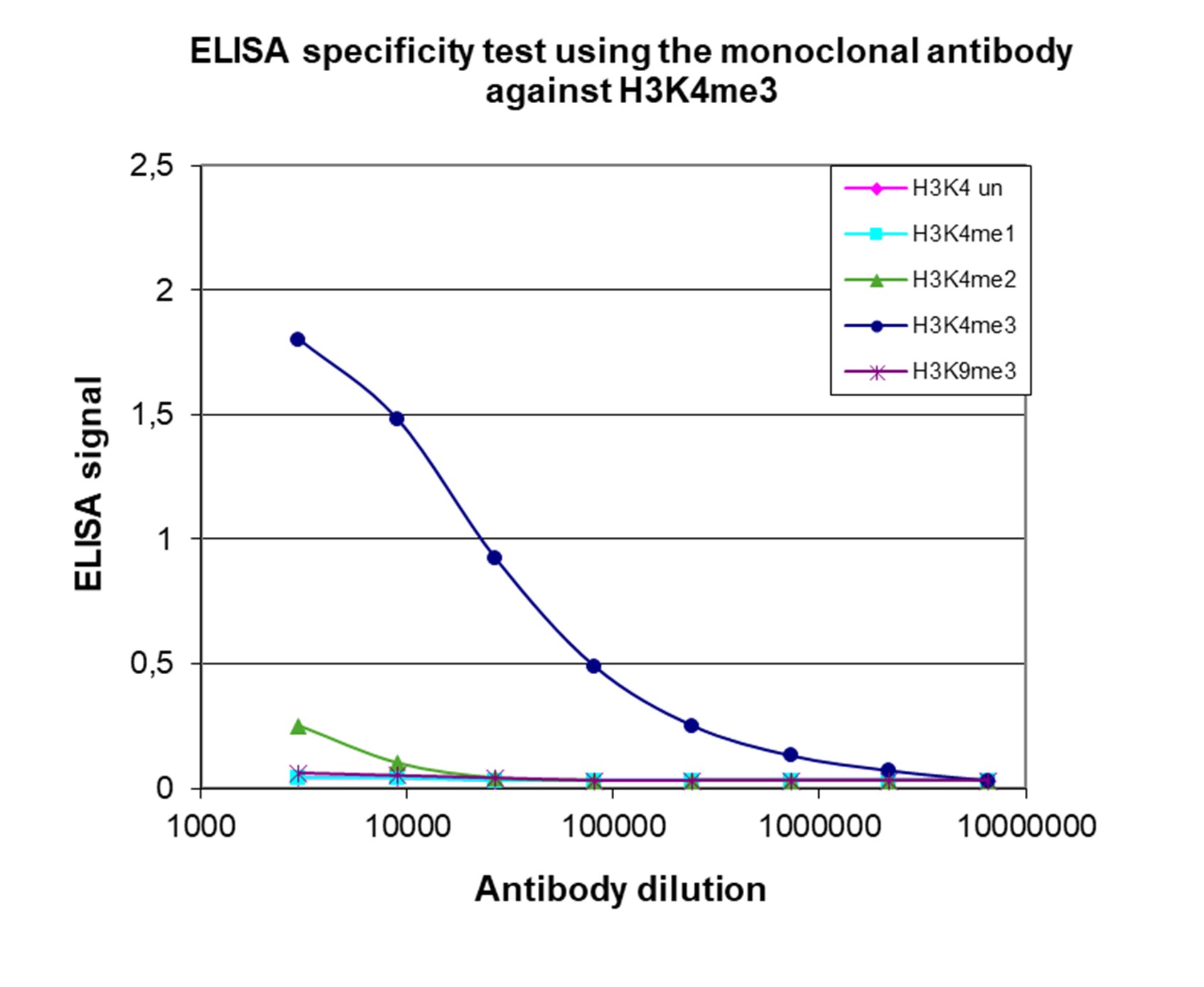
Figure 4. Cross-reactivity of the monoclonal antibody directed against H3K4me3
To test the specificity, an ELISA was performed using a serial dilution of the monoclonal antibody against H3K4me3 (cat. no. C15200152). The wells were coated with peptides containing the unmodified H3K4 as well as the mono-, di, and trimethylated H3K4 and the trimethylated H3K9. Figure 4 shows a high specificity of the antibody for the modification of interest.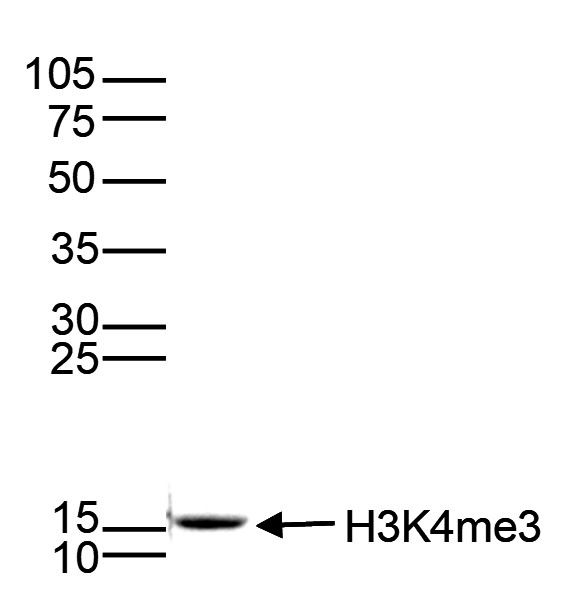
Figure 5. Western blot analysis using the monoclonal antibody directed against H3K4me3
Histone extracts (15 μg) from HeLa cells were analyzed by Western blot using the monoclonal antibody against H3K4me3 (cat. no. C15200152) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left.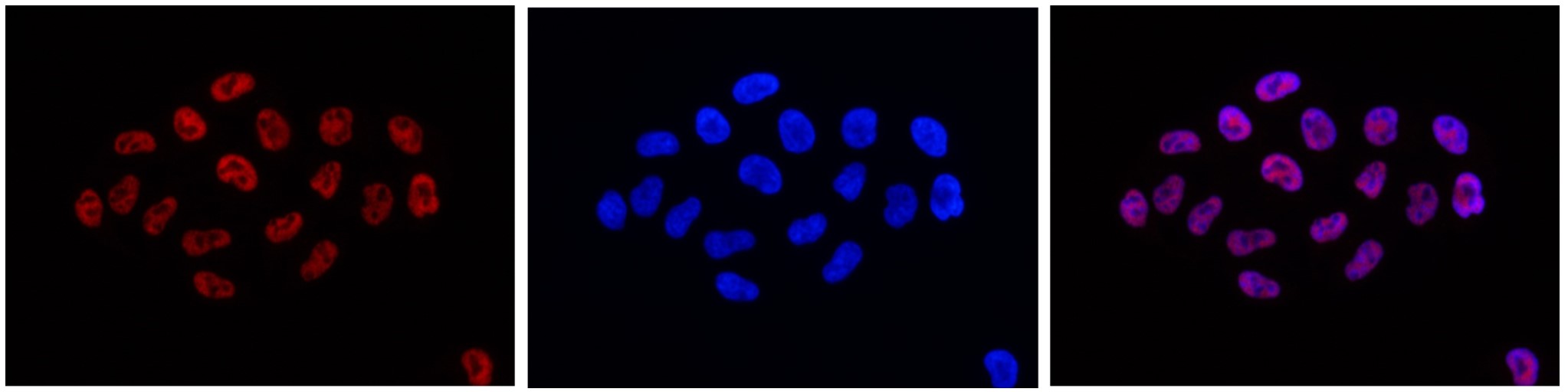
Figure 6. Immunofluorescence using the monoclonal antibody directed against H3K4me3
HeLa cells were stained with the antibody against H3K4me3 (cat. no. C15200152) and with DAPI. Cells were fixed with 4% formaldehyde for 10** min** and blocked with PBS/TX-100 containing 5% normal goat serum and 1% BSA. The cells were immunofluorescently labeled with the H3K4me3 antibody (left) diluted 1:500 in blocking solution, followed by an anti-mouse antibody conjugated to Alexa 594. The middle panel shows staining of the nuclei with DAPI. A merge of the two stains is shown on the right. - Publications

How to properly cite our product/service in your work
We strongly recommend using this: H3K4me3 Antibody (Hologic Diagenode Cat# C15200152 Lot# 001-19). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
The brain neurovascular epigenome and its association with dementia
Ziegler, Kevin Chris et al.
Cerebral small vessel disease (SVD) is frequently comorbid with Alzheimer's disease (AD), and brain endothelial cells (BECs) express genes associated with AD genetic risk. However, the epigenome of neurovascular cells and its intersection with genetic risk remain unexplored. Here, we generated gene regulomes for h...Microbiota translocation following intestinal barrier disruption promotes Mincle-mediated training of myeloid progenitors in the bone marrow
Iñaki Robles-Vera et al.
Highlights Gut barrier disruption induces trained immunity in bone marrow progenitors Enterococcus faecalis translocates to the bone marrow, inducing trained immunity Mincle sensing of E. faecalis mediates trained immunity of bone marrow progenitors Mincle-me...Local euchromatin enrichment in lamina-associated domains anticipatestheir repositioning in the adipogenic lineage.
Madsen-Østerbye J. et al.
BACKGROUND: Interactions of chromatin with the nuclear lamina via lamina-associated domains (LADs) confer structural stability to the genome. The dynamics of positioning of LADs during differentiation, and how LADs impinge on developmental gene expression, remains, however, elusive. RESULTS: We examined changes in t...The glucocorticoid receptor recruits the COMPASS complex to regulateinflammatory transcription at macrophage enhancers.
Greulich, Franziska et al.
Glucocorticoids (GCs) are effective anti-inflammatory drugs; yet, their mechanisms of action are poorly understood. GCs bind to the glucocorticoid receptor (GR), a ligand-gated transcription factor controlling gene expression in numerous cell types. Here, we characterize GR's protein interactome and find the SETD1A ...Distinct and temporary-restricted epigenetic mechanisms regulate human αβ and γδ T cell development
Roels J, Kuchmiy A, De Decker M, et al.
The development of TCRαβ and TCRγδ T cells comprises a step-wise process in which regulatory events control differentiation and lineage outcome. To clarify these mechanisms, we employed RNA-sequencing, ATAC-sequencing and ChIPmentation on well-defined thymocyte subsets that represent the conti...High resolution methylation analysis of the HoxA5 regulatory region in different somatic tissues of laboratory mouse during development
Sinha P. et al.
Homeobox genes encode a group of DNA binding regulatory proteins whose key function occurs in the spatial-temporal organization of genome during embryonic development and differentiation. The role of these Hox genes during ontogenesis makes it an important model for research. HoxA5 is a member of Hox gene family pla...Detection of Protein-Protein Interactions and Posttranslational Modifications Using the Proximity Ligation Assay: Application to the Study of the SUMO Pathway.
Ristic M. et al.
The detection of protein-protein interactions by imaging techniques often requires the overexpression of the proteins of interest tagged with fluorescent molecules, which can affect their biological properties and, subsequently, flaw experiment interpretations. The recent development of the proximity ligation assays...Methylation of the Sox9 and Oct4 promoters and its correlation with gene expression during testicular development in the laboratory mouse
Pamnani M et al.
Sox9 and Oct4 are two important regulatory factors involved in mammalian development. Sox9, a member of the group E Sox transcription factor family, has a crucial role in the development of the genitourinary system, while Oct4, commonly known as octamer binding transcription factor 4, belongs to class V of the trans...Germline organization in Strongyloides nematodes reveals alternative differentiation and regulation mechanisms.
Kulkarni A et al.
Nematodes of the genus Strongyloides are important parasites of vertebrates including man. Currently, little is known about their germline organization or reproductive biology and how this influences their parasitic life strategies. Here, we analyze the structure of the germline in several Strongyloides and closely ...Transcription termination and chimeric RNA formation controlled by Arabidopsis thaliana FPA.
Duc C, Sherstnev A, Cole C, Barton GJ, Simpson GG
Alternative cleavage and polyadenylation influence the coding and regulatory potential of mRNAs and where transcription termination occurs. Although widespread, few regulators of this process are known. The Arabidopsis thaliana protein FPA is a rare example of a trans-acting regulator of poly(A) site choice. Analysi...