Notice (8): Undefined variable: solution_of_interest [APP/View/Products/view.ctp, line 755]Code Context<!-- BEGIN: REQUEST_FORM MODAL -->
<div id="request_formModal" class="reveal-modal medium" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<?= $this->element('Forms/simple_form', array('solution_of_interest' => $solution_of_interest, 'header' => $header, 'message' => $message, 'campaign_id' => $campaign_id)) ?>
$viewFile = '/home/website-server/www/app/View/Products/view.ctp'
$dataForView = array(
'language' => 'en',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'meta_title' => 'MicroChIP DiaPure columns',
'product' => array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
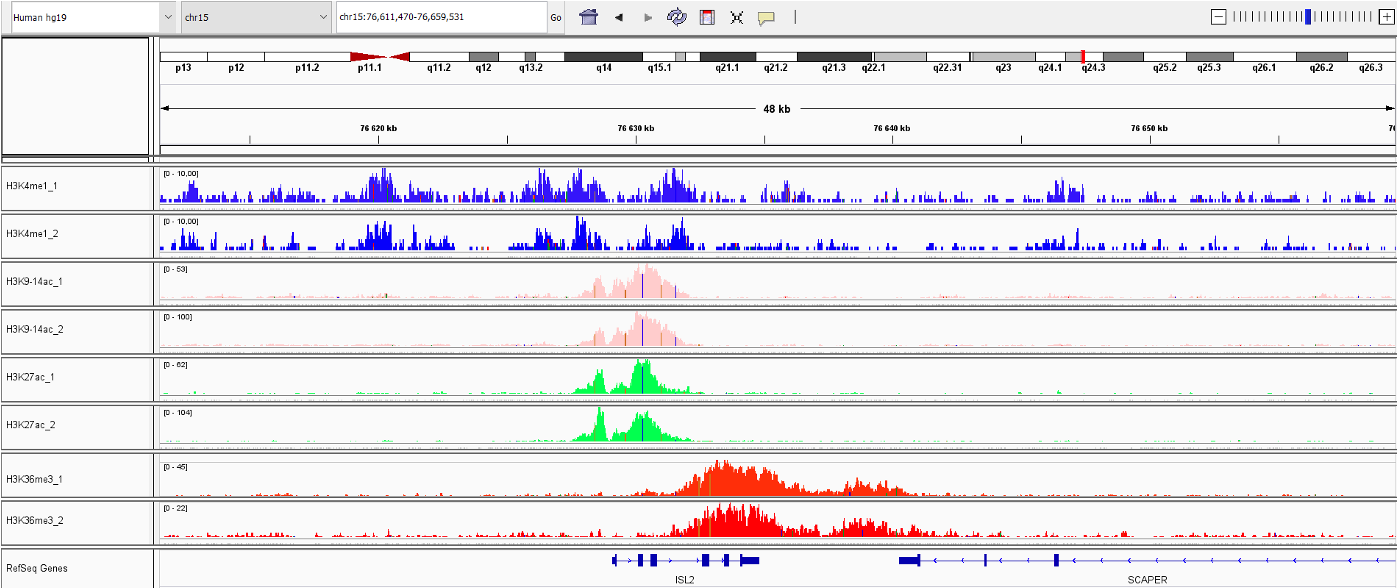
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
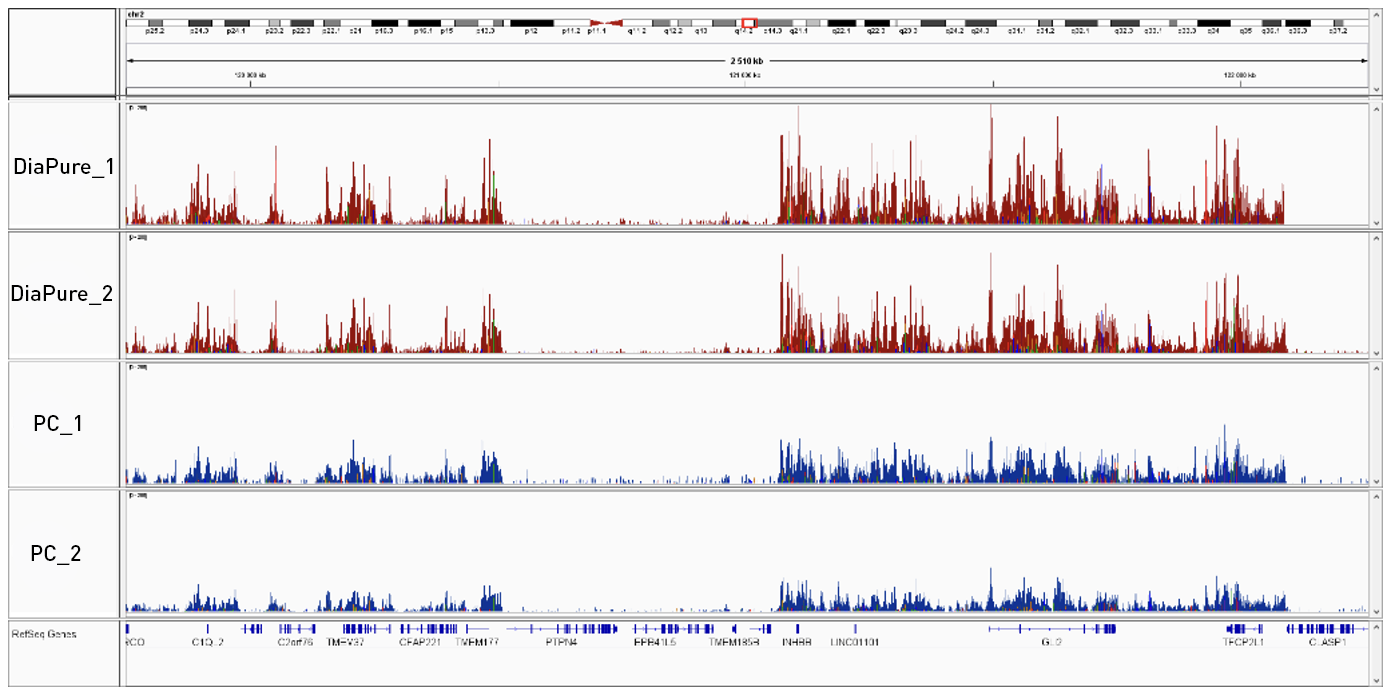
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Application' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
)
),
'Category' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Document' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
),
(int) 8 => array(
[maximum depth reached]
),
(int) 9 => array(
[maximum depth reached]
),
(int) 10 => array(
[maximum depth reached]
),
(int) 11 => array(
[maximum depth reached]
),
(int) 12 => array(
[maximum depth reached]
),
(int) 13 => array(
[maximum depth reached]
),
(int) 14 => array(
[maximum depth reached]
),
(int) 15 => array(
[maximum depth reached]
),
(int) 16 => array(
[maximum depth reached]
),
(int) 17 => array(
[maximum depth reached]
),
(int) 18 => array(
[maximum depth reached]
),
(int) 19 => array(
[maximum depth reached]
),
(int) 20 => array(
[maximum depth reached]
),
(int) 21 => array(
[maximum depth reached]
),
(int) 22 => array(
[maximum depth reached]
),
(int) 23 => array(
[maximum depth reached]
),
(int) 24 => array(
[maximum depth reached]
),
(int) 25 => array(
[maximum depth reached]
),
(int) 26 => array(
[maximum depth reached]
),
(int) 27 => array(
[maximum depth reached]
),
(int) 28 => array(
[maximum depth reached]
),
(int) 29 => array(
[maximum depth reached]
),
(int) 30 => array(
[maximum depth reached]
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
)
)
)
)
$language = 'en'
$meta_keywords = ''
$meta_description = 'MicroChIP DiaPure columns'
$meta_title = 'MicroChIP DiaPure columns'
$product = array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
[maximum depth reached]
),
'Image' => array(
[maximum depth reached]
)
)
),
'Application' => array(
(int) 0 => array(
'id' => '10',
'position' => '10',
'parent_id' => '2',
'name' => 'ChIP-qPCR',
'description' => '<div class="row">
<div class="small-12 medium-12 large-12 columns text-justify">
<p class="text-justify">Chromatin Immunoprecipitation (ChIP) coupled with quantitative PCR can be used to investigate protein-DNA interaction at known genomic binding sites. if sites are not known, qPCR primers can also be designed against potential regulatory regions such as promoters. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of performing real-time PCR is minimal. This technique is now used in a variety of life science disciplines including cellular differentiation, tumor suppressor gene silencing, and the effect of histone modifications on gene expression.</p>
<p class="text-justify"><strong>The ChIP-qPCR workflow</strong></p>
</div>
<div class="small-12 medium-12 large-12 columns text-center"><br /> <img src="https://www.diagenode.com/img/chip-qpcr-diagram.png" /></div>
<div class="small-12 medium-12 large-12 columns"><br />
<ol>
<li class="large-12 columns"><strong>Chromatin preparation: </strong>cell fixation (cross-linking) of chromatin-bound proteins such as histones or transcription factors to DNA followed by cell lysis.</li>
<li class="large-12 columns"><strong>Chromatin shearing: </strong>fragmentation of chromatin<strong> </strong>by sonication down to desired fragment size (100-500 bp)</li>
<li class="large-12 columns"><strong>Chromatin IP</strong>: protein-DNA complexe capture using<strong> <a href="https://www.diagenode.com/en/categories/chip-grade-antibodies">specific ChIP-grade antibodies</a></strong> against the histone or transcription factor of interest</li>
<li class="large-12 columns"><strong>DNA purification</strong>: chromatin reverse cross-linking and elution followed by purification<strong> </strong></li>
<li class="large-12 columns"><strong>qPCR and analysis</strong>: using previously designed primers to amplify IP'd material at specific loci</li>
</ol>
</div>
</div>
<div class="row" style="margin-top: 32px;">
<div class="small-12 medium-10 large-9 small-centered columns">
<div class="radius panel" style="background-color: #fff;">
<h3 class="text-center" style="color: #b21329;">Need guidance?</h3>
<p class="text-justify">Choose our full ChIP kits or simply choose what you need from antibodies, buffers, beads, chromatin shearing and purification reagents. With the ChIP Kit Customizer, you have complete flexibility on which components you want from our validated ChIP kits.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/which-kit-to-choose"><img src="https://www.diagenode.com/img/banners/banner-decide.png" alt="" /></a></div>
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/chip-kit-customizer-1"><img src="https://www.diagenode.com/img/banners/banner-customizer.png" alt="" /></a></div>
</div>
</div>
</div>
</div>',
'in_footer' => false,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'chip-qpcr',
'meta_keywords' => 'Chromatin immunoprecipitation,ChIP Quantitative PCR,polymerase chain reaction (PCR)',
'meta_description' => 'Diagenode's ChIP qPCR kits can be used to quantify enriched DNA after chromatin immunoprecipitation. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of',
'meta_title' => 'ChIP Quantitative PCR (ChIP-qPCR) | Diagenode',
'modified' => '2018-01-09 16:46:56',
'created' => '2014-12-11 00:22:08',
'ProductsApplication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '2',
'position' => '10',
'parent_id' => null,
'name' => 'エピジェネティクス・クロマチン解析',
'description' => '<div class="row">
<div class="large-12 columns">エピジェネティクス研究は、異なる転写パターン、遺伝子発現およびサイレンシングを引き起こすクロマチンの変化に対処します。<br /><br />クロマチンの主成分はDNA<span>およびヒストン蛋白質です。<span> </span></span>各ヒストンコア蛋白質(H2A<span>、</span>H2B<span>、</span>H3<span>および</span>H4<span>)の</span>2<span>つのコピーを</span>8<span>量体に組み込み、</span>DNA<span>で包んでヌクレオソームコアを形成させます。<span> </span></span>ヌクレオソームは、転写機械のDNA<span>への接近可能性および</span>クロマチン再構成因子を制御します。</div>
<div class="large-12 columns">
<p></p>
<p>クロマチン免疫沈降(ChIP<span>)は、関心対象の特定の蛋白質に対するゲノム結合部位の位置を解明するために使用される方法であり、遺伝子発現の制御に関する非常に貴重な洞察を提供します。<span> </span></span>ChIPは特定の抗原を含むクロマチン断片の選択的富化に関与します。 特定の蛋白質または蛋白質修飾を認識する抗体を使用して、特定の遺伝子座における抗原の相対存在量を決定します。</p>
<p>ChIP-seq<span>および</span>ChIP-qPCR<span>は、蛋白質</span>-DNA<span>結合部位の同定を可能にする技術です。</span></p>
<p> </p>
</div>
</div>',
'in_footer' => true,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'epigenetics-chromatin-study',
'meta_keywords' => 'Chromatin Immunoprecipitation Sequencing,ChIP-Seq,ChIP-seq grade antibodies,DNA purification,qPCR,Shearing of chromatin',
'meta_description' => 'Diagenodeは、ChIP-Seqおよびクロマチン免疫沈降(ChIP)専用キットと抗体を定量PCRアプリケーションと組み合わせて幅広く提供しています。',
'meta_title' => 'エピジェネティクス - クロマチン免疫沈降(ChIP)| Diagenode',
'modified' => '2018-01-11 09:58:47',
'created' => '2015-02-20 16:10:25',
'ProductsApplication' => array(
[maximum depth reached]
)
)
),
'Category' => array(
(int) 0 => array(
'id' => '61',
'position' => '2',
'parent_id' => '14',
'name' => 'Column-based DNA purification',
'description' => '',
'no_promo' => false,
'in_menu' => false,
'online' => true,
'tabular' => true,
'hide' => false,
'all_format' => false,
'is_antibody' => false,
'slug' => 'column-based-dna-purification',
'cookies_tag_id' => null,
'meta_keywords' => 'MicroChIP DiaPure columns,diagenode',
'meta_description' => 'Diagenode provides Column-based DNA purification for downstream analysis.',
'meta_title' => 'Column-based DNA purification | Diagenode',
'modified' => '2016-01-21 13:49:21',
'created' => '2015-07-08 10:31:04',
'ProductsCategory' => array(
[maximum depth reached]
),
'CookiesTag' => array([maximum depth reached])
)
),
'Document' => array(
(int) 0 => array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
[maximum depth reached]
)
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
'id' => '5033',
'name' => 'Trithorax regulates long-term memory in Drosophila through epigenetic maintenance of mushroom body metabolic state and translation capacity',
'authors' => 'Nicholas Raun et al.',
'description' => '<p><span>The role of epigenetics and chromatin in the maintenance of postmitotic neuronal cell identities is not well understood. Here, we show that the histone methyltransferase Trithorax (Trx) is required in postmitotic memory neurons of the </span><em>Drosophila</em><span><span> </span>mushroom body (MB) to enable their capacity for long-term memory (LTM), but not short-term memory (STM). Using MB-specific RNA-, ChIP-, and ATAC-sequencing, we find that Trx maintains homeostatic expression of several non-canonical MB-enriched transcripts, including the orphan nuclear receptor<span> </span></span><em>Hr51</em><span>, and the metabolic enzyme<span> </span></span><em>lactate dehydrogenase</em><span><span> </span>(</span><em>Ldh)</em><span>. Through these key targets, Trx facilitates a metabolic state characterized by high lactate levels in MBγ neurons. This metabolic state supports a high capacity for protein translation, a process that is essential for LTM, but not STM. These data suggest that Trx, a classic regulator of cell type specification during development, has additional functions in maintaining underappreciated aspects of postmitotic neuron identity, such as metabolic state. Our work supports a body of evidence suggesting that a high capacity for energy metabolism is an essential cell identity characteristic for neurons that mediate LTM.</span></p>',
'date' => '2025-01-27',
'pmid' => 'https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.3003004',
'doi' => 'https://doi.org/10.1371/journal.pbio.3003004',
'modified' => '2025-01-30 10:12:42',
'created' => '2025-01-30 10:12:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '4983',
'name' => 'Integrated multi-omics analysis of PBX1 in mouse adult neural stem- and progenitor cells identifies a transcriptional module that functionally links PBX1 to TCF3/4',
'authors' => 'Vera Laub et al.',
'description' => '<p><span>Developmental transcription factors act in networks, but how these networks achieve cell- and tissue specificity is still poorly understood. Here, we explored pre-B cell leukemia homeobox 1 (PBX1) in adult neurogenesis combining genomic, transcriptomic, and proteomic approaches. ChIP-seq analysis uncovered PBX1 binding to numerous genomic sites. Integration of PBX1 ChIP-seq with ATAC-seq data predicted interaction partners, which were subsequently validated by mass spectrometry. Whole transcriptome spatial RNA analysis revealed shared expression dynamics of </span><em>Pbx1</em><span><span> </span>and interacting factors. Among these were class I bHLH proteins TCF3 and TCF4. RNA-seq following<span> </span></span><em>Pbx1</em><span>,<span> </span></span><em>Tcf3</em><span><span> </span>or<span> </span></span><em>Tcf4</em><span><span> </span>knockdown identified proliferation- and differentiation associated genes as shared targets, while sphere formation assays following knockdown argued for functional cooperativity of PBX1 and TCF3 in progenitor cell proliferation. Notably, while physiological PBX1-TCF interaction has not yet been described, chromosomal translocation resulting in genomic<span> </span></span><em>TCF3::PBX1</em><span><span> </span>fusion characterizes a subtype of acute lymphoblastic leukemia. Introducing<span> </span></span><em>Pbx1</em><span><span> </span>into Nalm6 cells, a pre-B cell line expressing<span> </span></span><em>TCF3</em><span><span> </span>but lacking<span> </span></span><em>PBX1</em><span>, upregulated the leukemogenic genes<span> </span></span><em>BLK</em><span><span> </span>and<span> </span></span><em>NOTCH3</em><span>, arguing that functional PBX1-TCF cooperativity likely extends to hematopoiesis. Our study hence uncovers a transcriptional module orchestrating the balance between progenitor cell proliferation and differentiation in adult neurogenesis with potential implications for leukemia etiology.</span></p>',
'date' => '2024-10-08',
'pmid' => 'https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkae864/7815639',
'doi' => 'https://doi.org/10.1093/nar/gkae864',
'modified' => '2024-10-11 10:02:42',
'created' => '2024-10-11 10:02:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '4964',
'name' => 'A PRE loop at the dac locus acts as a topological chromatin structure that restricts and specifies enhancer–promoter communication',
'authors' => 'Denaud S. et al.',
'description' => '<p><span>Three-dimensional (3D) genome folding has a fundamental role in the regulation of developmental genes by facilitating or constraining chromatin interactions between </span><i>cis</i><span>-regulatory elements (CREs). Polycomb response elements (PREs) are a specific kind of CRE involved in the memory of transcriptional states in<span> </span></span><i>Drosophila melanogaster</i><span>. PREs act as nucleation sites for Polycomb group (PcG) proteins, which deposit the repressive histone mark H3K27me3, leading to the formation of a class of topologically associating domain (TAD) called a Polycomb domain. PREs can establish looping contacts that stabilize the gene repression of key developmental genes during development. However, the mechanism by which PRE loops fine-tune gene expression is unknown. Using clustered regularly interspaced short palindromic repeats and Cas9 genome engineering, we specifically perturbed PRE contacts or enhancer function and used complementary approaches including 4C-seq, Hi-C and Hi-M to analyze how chromatin architecture perturbation affects gene expression. Our results suggest that the PRE loop at the<span> </span></span><i>dac</i><span><span> </span>gene locus acts as a constitutive 3D chromatin scaffold during<span> </span></span><i>Drosophila</i><span><span> </span>development that forms independently of gene expression states and has a versatile function; it restricts enhancer–promoter communication and contributes to enhancer specificity.</span></p>',
'date' => '2024-08-16',
'pmid' => 'https://www.nature.com/articles/s41594-024-01375-7',
'doi' => 'https://doi.org/10.1038/s41594-024-01375-7',
'modified' => '2024-09-02 10:20:28',
'created' => '2024-09-02 10:20:28',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '4954',
'name' => 'A multiomic atlas of the aging hippocampus reveals molecular changes in response to environmental enrichment',
'authors' => 'Perez R. F. at al. ',
'description' => '<p><span>Aging involves the deterioration of organismal function, leading to the emergence of multiple pathologies. Environmental stimuli, including lifestyle, can influence the trajectory of this process and may be used as tools in the pursuit of healthy aging. To evaluate the role of epigenetic mechanisms in this context, we have generated bulk tissue and single cell multi-omic maps of the male mouse dorsal hippocampus in young and old animals exposed to environmental stimulation in the form of enriched environments. We present a molecular atlas of the aging process, highlighting two distinct axes, related to inflammation and to the dysregulation of mRNA metabolism, at the functional RNA and protein level. Additionally, we report the alteration of heterochromatin domains, including the loss of bivalent chromatin and the uncovering of a heterochromatin-switch phenomenon whereby constitutive heterochromatin loss is partially mitigated through gains in facultative heterochromatin. Notably, we observed the multi-omic reversal of a great number of aging-associated alterations in the context of environmental enrichment, which was particularly linked to glial and oligodendrocyte pathways. In conclusion, our work describes the epigenomic landscape of environmental stimulation in the context of aging and reveals how lifestyle intervention can lead to the multi-layered reversal of aging-associated decline.</span></p>',
'date' => '2024-07-16',
'pmid' => 'https://www.nature.com/articles/s41467-024-49608-z',
'doi' => 'https://doi.org/10.1038/s41467-024-49608-z',
'modified' => '2024-07-29 11:33:49',
'created' => '2024-07-29 11:33:49',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '4927',
'name' => 'Inflammatory stress-mediated chromatin changes underlie dysfunction in endothelial cells',
'authors' => 'Liu H. et al.',
'description' => '<p><span>Inflammatory stresses underlie endothelial dysfunction and contribute to the development of chronic cardiovascular disorders such as atherosclerosis and vascular fibrosis. The initial transcriptional response of endothelial cells to pro-inflammatory cytokines such as TNF-alpha is well established. However, very few studies uncover the effects of inflammatory stresses on chromatin architecture. We used integrative analysis of ATAC-seq and RNA-seq data to investigate chromatin alterations in human endothelial cells in response to TNF-alpha and febrile-range heat stress exposure. Multi-omics data analysis suggests a correlation between the transcription of stress-related genes and endothelial dysfunction drivers with chromatin regions exhibiting differential accessibility. Moreover, microscopy identified the dynamics in the nuclear organization, specifically, the changes in a subset of heterochromatic nucleoli-associated chromatin domains, the centromeres. Upon inflammatory stress exposure, the centromeres decreased association with nucleoli in a p38-dependent manner and increased the number of transcripts from pericentromeric regions. Overall, we provide two lines of evidence that suggest chromatin alterations in vascular endothelial cells during inflammatory stresses.</span></p>',
'date' => '2023-10-16',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10614786/',
'doi' => '10.1101/2023.10.11.561959',
'modified' => '2024-03-27 15:15:21',
'created' => '2024-03-27 15:15:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '4862',
'name' => 'Mutant FUS induces chromatin reorganization in the hippocampus andalters memory processes.',
'authors' => 'Tzeplaeff L. et al.',
'description' => '<p>Cytoplasmic mislocalization of the nuclear Fused in Sarcoma (FUS) protein is associated to amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Cytoplasmic FUS accumulation is recapitulated in the frontal cortex and spinal cord of heterozygous Fus mice. Yet, the mechanisms linking FUS mislocalization to hippocampal function and memory formation are still not characterized. Herein, we show that in these mice, the hippocampus paradoxically displays nuclear FUS accumulation. Multi-omic analyses showed that FUS binds to a set of genes characterized by the presence of an ETS/ELK-binding motifs, and involved in RNA metabolism, transcription, ribosome/mitochondria and chromatin organization. Importantly, hippocampal nuclei showed a decompaction of the neuronal chromatin at highly expressed genes and an inappropriate transcriptomic response was observed after spatial training of Fus mice. Furthermore, these mice lacked precision in a hippocampal-dependent spatial memory task and displayed decreased dendritic spine density. These studies shows that mutated FUS affects epigenetic regulation of the chromatin landscape in hippocampal neurons, which could participate in FTD/ALS pathogenic events. These data call for further investigation in the neurological phenotype of FUS-related diseases and open therapeutic strategies towards epigenetic drugs.</p>',
'date' => '2023-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/37327984',
'doi' => '10.1016/j.pneurobio.2023.102483',
'modified' => '2023-08-01 14:55:49',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '4733',
'name' => 'FXR inhibition may protect from SARS-CoV-2 infection by reducingACE2.',
'authors' => 'Brevini Teresa et al.',
'description' => '<p>Prevention of SARS-CoV-2 infection through the modulation of viral host receptors, such as ACE2, could represent a new chemoprophylactic approach for COVID-19 complementing vaccination. However, the mechanisms controlling ACE2 expression remain elusive. Here, we identify the farnesoid X receptor (FXR) as a direct regulator of ACE2 transcription in multiple COVID19-affected tissues, including the gastrointestinal and respiratory systems. We then use the over-the-counter compound z-guggulsterone (ZGG) and the off-patent drug ursodeoxycholic acid (UDCA) to reduce FXR signalling and downregulate ACE2 in human lung, cholangiocyte and intestinal organoids and in the corresponding tissues in mice and hamsters. We demonstrate that UDCA-mediated ACE2 downregulation reduces susceptibility to SARS-CoV-2 infection in vitro, in vivo and in human lungs and livers perfused ex situ. Furthermore, we illustrate that UDCA reduces ACE2 expression in the nasal epithelium in humans. Finally, we identify a correlation between UDCA treatment and positive clinical outcomes following SARS-CoV-2 infection using retrospective registry data, and confirm these findings in an independent validation cohort of liver transplant recipients. In conclusion, we identify a novel function of FXR in controlling ACE2 expression and provide evidence that modulation of this pathway could be beneficial for reducing SARS-CoV-2 infection, paving the road for future clinical trials.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36470304',
'doi' => '10.1038/s41586-022-05594-0',
'modified' => '2023-03-13 08:52:11',
'created' => '2023-02-28 12:19:11',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '4575',
'name' => 'Intranasal administration of Acinetobacter lwoffii in a murine model ofasthma induces IL-6-mediated protection associated with cecal microbiotachanges.',
'authors' => 'Alashkar A. B. et al.',
'description' => '<p>BACKGROUND: Early-life exposure to certain environmental bacteria including Acinetobacter lwoffii (AL) has been implicated in protection from chronic inflammatory diseases including asthma later in life. However, the underlying mechanisms at the immune-microbe interface remain largely unknown. METHODS: The effects of repeated intranasal AL exposure on local and systemic innate immune responses were investigated in wild-type and Il6 , Il10 , and Il17 mice exposed to ovalbumin-induced allergic airway inflammation. Those investigations were expanded by microbiome analyses. To assess for AL-associated changes in gene expression, the picture arising from animal data was supplemented by in vitro experiments of macrophage and T-cell responses, yielding expression and epigenetic data. RESULTS: The asthma preventive effect of AL was confirmed in the lung. Repeated intranasal AL administration triggered a proinflammatory immune response particularly characterized by elevated levels of IL-6, and consequently, IL-6 induced IL-10 production in CD4 T-cells. Both IL-6 and IL-10, but not IL-17, were required for asthma protection. AL had a profound impact on the gene regulatory landscape of CD4 T-cells which could be largely recapitulated by recombinant IL-6. AL administration also induced marked changes in the gastrointestinal microbiome but not in the lung microbiome. By comparing the effects on the microbiota according to mouse genotype and AL-treatment status, we have identified microbial taxa that were associated with either disease protection or activity. CONCLUSION: These experiments provide a novel mechanism of Acinetobacter lwoffii-induced asthma protection operating through IL-6-mediated epigenetic activation of IL-10 production and with associated effects on the intestinal microbiome.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36458896',
'doi' => '10.1111/all.15606',
'modified' => '2023-04-11 10:23:07',
'created' => '2023-02-21 09:59:46',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 8 => array(
'id' => '4836',
'name' => 'Caffeine intake exerts dual genome-wide effects on hippocampal metabolismand learning-dependent transcription.',
'authors' => 'Paiva I. et al.',
'description' => '<p>Caffeine is the most widely consumed psychoactive substance in the world. Strikingly, the molecular pathways engaged by its regular consumption remain unclear. We herein addressed the mechanisms associated with habitual (chronic) caffeine consumption in the mouse hippocampus using untargeted orthogonal omics techniques. Our results revealed that chronic caffeine exerts concerted pleiotropic effects in the hippocampus at the epigenomic, proteomic, and metabolomic levels. Caffeine lowered metabolism-related processes (e.g., at the level of metabolomics and gene expression) in bulk tissue, while it induced neuron-specific epigenetic changes at synaptic transmission/plasticity-related genes and increased experience-driven transcriptional activity. Altogether, these findings suggest that regular caffeine intake improves the signal-to-noise ratio during information encoding, in part through fine-tuning of metabolic genes, while boosting the salience of information processing during learning in neuronal circuits.</p>',
'date' => '2022-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35536645',
'doi' => '10.1172/JCI149371',
'modified' => '2023-08-01 13:52:29',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 9 => array(
'id' => '4220',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in Mouse Prostate Cancer Xenografts',
'authors' => 'Sanchez A. et al.',
'description' => '<p><strong class="sub-title">Background/aim:<span> </span></strong>Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo.</p>
<p><strong class="sub-title">Materials and methods:<span> </span></strong>Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR.</p>
<p><strong class="sub-title">Results:<span> </span></strong>JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression.</p>
<p><strong class="sub-title">Conclusion:<span> </span></strong>JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-05-01',
'pmid' => 'https://pubmed.ncbi.nlm.nih.gov/35430567/',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-04-21 11:54:21',
'created' => '2022-04-21 11:54:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 10 => array(
'id' => '4409',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in MouseProstate Cancer Xenografts.',
'authors' => 'Sanchez A. et al.',
'description' => '<p>BACKGROUND/AIM: Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo. MATERIALS AND METHODS: Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR. RESULTS: JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression. CONCLUSION: JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35430567',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-08-11 15:11:58',
'created' => '2022-08-11 12:14:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 11 => array(
'id' => '4233',
'name' => 'Autocrine Vitamin D-signaling switches off pro-inflammatory programsof Th1 cells',
'authors' => 'Chauss D.et al.',
'description' => '<p>The molecular mechanisms governing orderly shutdown and retraction of CD4+ T helper (Th)1 responses remain poorly understood. Here, we show that complement triggers contraction of Th1 responses by inducing intrinsic expression of the vitamin D (VitD) receptor (VDR) and the VitD-activating enzyme CYP27B1, permitting T cells to both activate and respond to VitD. VitD then initiated transition from pro-inflammatory IFN-γ + Th1 cells to suppressive IL-10+ cells. This process was primed by dynamic changes in the epigenetic landscape of CD4+ T cells, generating super-enhancers and recruiting several transcription factors, notably c-JUN, STAT3 and BACH2, which together with VDR shaped the transcriptional response to VitD. Accordingly, VitD did not induce IL-10 in cells with dysfunctional BACH2 or STAT3. Bronchoalveolar lavage fluid CD4+ T cells of COVID-19 patients were Th1-skewed and showed de-repression of genes down-regulated by VitD, either from lack of substrate (VitD deficiency) and/or abnormal regulation of this system.</p>',
'date' => '2021-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/34764490',
'doi' => '10.1038/s41590-021-01080-3',
'modified' => '2022-05-19 16:57:27',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 12 => array(
'id' => '4351',
'name' => 'Essential role of a ThPOK autoregulatory loop in the maintenance ofmature CD4 T cell identity and function.',
'authors' => 'Basu Jayati et al.',
'description' => '<p>The transcription factor ThPOK (encoded by the Zbtb7b gene) controls homeostasis and differentiation of mature helper T cells, while opposing their differentiation to CD4 intraepithelial lymphocytes (IELs) in the intestinal mucosa. Thus CD4 IEL differentiation requires ThPOK transcriptional repression via reactivation of the ThPOK transcriptional silencer element (Sil). In the present study, we describe a new autoregulatory loop whereby ThPOK binds to the Sil to maintain its own long-term expression in CD4 T cells. Disruption of this loop in vivo prevents persistent ThPOK expression, leads to genome-wide changes in chromatin accessibility and derepresses the colonic regulatory T (T) cell gene expression signature. This promotes selective differentiation of naive CD4 T cells into GITRPD-1CD25 (Triple) T cells and conversion to CD4 IELs in the gut, thereby providing dominant protection from colitis. Hence, the ThPOK autoregulatory loop represents a key mechanism to physiologically control ThPOK expression and T cell differentiation in the gut, with potential therapeutic relevance.</p>',
'date' => '2021-08-01',
'pmid' => 'https://doi.org/10.1038%2Fs41590-021-00980-8',
'doi' => '10.1038/s41590-021-00980-8',
'modified' => '2022-06-22 12:32:59',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 13 => array(
'id' => '4127',
'name' => 'The histone modification H3K4me3 is altered at the locus in Alzheimer'sdisease brain.',
'authors' => 'Smith, Adam et al.',
'description' => '<p>Several epigenome-wide association studies of DNA methylation have highlighted altered DNA methylation in the gene in Alzheimer's disease (AD) brain samples. However, no study has specifically examined histone modifications in the disease. We use chromatin immunoprecipitation-qPCR to quantify tri-methylation at histone 3 lysine 4 (H3K4me3) and 27 (H3K27me3) in the gene in entorhinal cortex from donors with high (n = 59) or low (n = 29) Alzheimer's disease pathology. We demonstrate decreased levels of H3K4me3, a marker of active gene transcription, with no change in H3K27me3, a marker of inactive genes. H3K4me3 is negatively correlated with DNA methylation in specific regions of the gene. Our study suggests that the gene shows altered epigenetic marks indicative of reduced gene activation in Alzheimer's disease.</p>',
'date' => '2021-02-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/33815817',
'doi' => '10.2144/fsoa-2020-0161',
'modified' => '2021-12-07 10:16:08',
'created' => '2021-12-06 15:53:19',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 14 => array(
'id' => '4062',
'name' => 'Digging Deeper into Breast Cancer Epigenetics: Insights from ChemicalInhibition of Histone Acetyltransferase TIP60 .',
'authors' => 'Idrissou, Mouhamed and Lebert, Andre and Boisnier, Tiphanie and Sanchez,Anna and Houfaf Khoufaf, Fatma Zohra and Penault-Llorca, Frédérique andBignon, Yves-Jean and Bernard-Gallon, Dominique',
'description' => '<p>Breast cancer is often sporadic due to several factors. Among them, the deregulation of epigenetic proteins may be involved. TIP60 or KAT5 is an acetyltransferase that regulates gene transcription through the chromatin structure. This pleiotropic protein acts in several cellular pathways by acetylating proteins. RNA and protein expressions of TIP60 were shown to decrease in some breast cancer subtypes, particularly in triple-negative breast cancer (TNBC), where a low expression of TIP60 was exhibited compared with luminal subtypes. In this study, the inhibition of the residual activity of TIP60 in breast cancer cell lines was investigated by using two chemical inhibitors, TH1834 and NU9056, first on the acetylation of the specific target, lysine 4 of histone 3 (H3K4) by immunoblotting, and second, by chromatin immunoprecipitation (ChIP)-qPCR (-quantitative Polymerase Chain Reaction). Subsequently, significant decreases or a trend toward decrease of H3K4ac in the different chromatin compartments were observed. In addition, the expression of 48 human nuclear receptors was studied with TaqMan Low-Density Array in these breast cancer cell lines treated with TIP60 inhibitors. The statistical analysis allowed us to comprehensively characterize the androgen receptor and receptors in TNBC cell lines after TH1834 or NU9056 treatment. The understanding of the residual activity of TIP60 in the evolution of breast cancer might be a major asset in the fight against this disease, and could allow TIP60 to be used as a biomarker or therapeutic target for breast cancer progression in the future.</p>',
'date' => '2020-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/32960142',
'doi' => '10.1089/omi.2020.0104',
'modified' => '2021-02-19 17:39:52',
'created' => '2021-02-18 10:21:53',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 15 => array(
'id' => '3969',
'name' => 'Delineating the early transcriptional specification of the mammalian trachea and esophagus.',
'authors' => 'Kuwahara A, Lewis AE, Coombes C, Leung FS, Percharde M, Bush JO',
'description' => '<p>The genome-scale transcriptional programs that specify the mammalian trachea and esophagus are unknown. Though NKX2-1 and SOX2 are hypothesized to be co-repressive master regulators of tracheoesophageal fates, this is untested at a whole transcriptomic scale and their downstream networks remain unidentified. By combining single-cell RNA-sequencing with bulk RNA-sequencing of mutants and NKX2-1 ChIP-sequencing in mouse embryos, we delineate the NKX2-1 transcriptional program in tracheoesophageal specification, and discover that the majority of the tracheal and esophageal transcriptome is NKX2-1 independent. To decouple the NKX2-1 transcriptional program from regulation by SOX2, we interrogate the expression of newly-identified tracheal and esophageal markers in / compound mutants. Finally, we discover that NKX2-1 binds directly to and and regulates their expression to control mesenchymal specification to cartilage and smooth muscle, coupling epithelial identity with mesenchymal specification. These findings create a new framework for understanding early tracheoesophageal fate specification at the genome-wide level.</p>',
'date' => '2020-06-09',
'pmid' => 'http://www.pubmed.gov/32515350',
'doi' => '10.7554/eLife.55526',
'modified' => '2020-08-12 09:32:02',
'created' => '2020-08-10 12:12:25',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 16 => array(
'id' => '3845',
'name' => 'Combinatorial action of NF-Y and TALE at embryonic enhancers defines distinct gene expression programs during zygotic genome activation in zebrafish.',
'authors' => 'Stanney W, Ladam F, Donaldson IJ, Parsons TJ, Maehr R, Bobola N, Sagerström CG',
'description' => '<p>Animal embryogenesis is initiated by maternal factors, but zygotic genome activation (ZGA) shifts regulatory control to the embryo during blastula stages. ZGA is thought to be mediated by maternally provided transcription factors (TFs), but few such TFs have been identified in vertebrates. Here we report that NF-Y and TALE TFs bind zebrafish genomic elements associated with developmental control genes already at ZGA. In particular, co-regulation by NF-Y and TALE is associated with broadly acting genes involved in transcriptional control, while regulation by either NF-Y or TALE defines genes in specific developmental processes, such that NF-Y controls a cilia gene expression program while TALE controls expression of hox genes. We also demonstrate that NF-Y and TALE-occupied genomic elements function as enhancers during embryogenesis. We conclude that combinatorial use of NF-Y and TALE at developmental enhancers permits the establishment of distinct gene expression programs at zebrafish ZGA.</p>',
'date' => '2019-12-17',
'pmid' => 'http://www.pubmed.gov/31862379',
'doi' => '10.1016/j.ydbio.2019.12.003',
'modified' => '2020-02-20 11:13:27',
'created' => '2020-02-13 10:02:44',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 17 => array(
'id' => '3744',
'name' => 'Whsc1 links pluripotency exit with mesendoderm specification.',
'authors' => 'Tian TV, Di Stefano B, Stik G, Vila-Casadesús M, Sardina JL, Vidal E, Dasti A, Segura-Morales C, De Andrés-Aguayo L, Gómez A, Goldmann J, Jaenisch R, Graf T',
'description' => '<p>How pluripotent stem cells differentiate into the main germ layers is a key question of developmental biology. Here, we show that the chromatin-related factor Whsc1 (also known as Nsd2 and MMSET) has a dual role in pluripotency exit and germ layer specification of embryonic stem cells. On induction of differentiation, a proportion of Whsc1-depleted embryonic stem cells remain entrapped in a pluripotent state and fail to form mesendoderm, although they are still capable of generating neuroectoderm. These functions of Whsc1 are independent of its methyltransferase activity. Whsc1 binds to enhancers of the mesendodermal regulators Gata4, T (Brachyury), Gata6 and Foxa2, together with Brd4, and activates the expression of these genes. Depleting each of these regulators also delays pluripotency exit, suggesting that they mediate the effects observed with Whsc1. Our data indicate that Whsc1 links silencing of the pluripotency regulatory network with activation of mesendoderm lineages.</p>',
'date' => '2019-07-01',
'pmid' => 'http://www.pubmed.gov/31235934',
'doi' => '10.1038/s41556-019-0342-1',
'modified' => '2019-08-06 16:35:35',
'created' => '2019-07-31 13:35:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 18 => array(
'id' => '3639',
'name' => 'Spatial confinement downsizes the inflammatory response of macrophages.',
'authors' => 'Jain N, Vogel V',
'description' => '<p>Macrophages respond to chemical/metabolic and physical stimuli, but their effects cannot be readily decoupled in vivo during pro-inflammatory activation. Here, we show that preventing macrophage spreading by spatial confinement, as imposed by micropatterning, microporous substrates or cell crowding, suppresses late lipopolysaccharide (LPS)-activated transcriptional programs (biomarkers IL-6, CXCL9, IL-1β, and iNOS) by mechanomodulating chromatin compaction and epigenetic alterations (HDAC3 levels and H3K36-dimethylation). Mechanistically, confinement reduces actin polymerization, thereby lowers the LPS-stimulated nuclear translocation of MRTF-A. This lowers the activity of the MRTF-A-SRF complex and subsequently downregulates the inflammatory response, as confirmed by chromatin immunoprecipitation coupled with quantitative PCR and RNA sequencing analysis. Confinement thus downregulates pro-inflammatory cytokine secretion and, well before any activation processes, the phagocytic potential of macrophages. Contrarily, early events, including activation of the LPS receptor TLR4, and downstream NF-κB and IRF3 signalling and hence the expression of early LPS-responsive genes were marginally affected by confinement. These findings have broad implications in the context of mechanobiology, inflammation and immunology, as well as in tissue engineering and regenerative medicine.</p>',
'date' => '2018-12-01',
'pmid' => 'http://www.pubmed.gov/30349032',
'doi' => '10.1038/s41563-018-0190-6',
'modified' => '2019-06-07 10:23:26',
'created' => '2019-06-06 12:11:18',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 19 => array(
'id' => '3379',
'name' => 'SIRT1-dependent epigenetic regulation of H3 and H4 histone acetylation in human breast cancer',
'authors' => 'Khaldoun Rifaï et al.',
'description' => '<p>Breast cancer is the most frequently diagnosed malignancy in women worldwide. It is well established that the complexity of carcinogenesis involves profound epigenetic deregulations that contribute to the tumorigenesis process. Deregulated H3 and H4 acetylated histone marks are amongst those alterations. Sirtuin-1 (SIRT1) is a class-III histone deacetylase deeply involved in apoptosis, genomic stability, gene expression regulation and breast tumorigenesis. However, the underlying molecular mechanism by which SIRT1 regulates H3 and H4 acetylated marks, and consequently cancer-related gene expression in breast cancer, remains uncharacterized. In this study, we elucidated SIRT1 epigenetic role and analyzed the link between the latter and histones H3 and H4 epigenetic marks in all 5 molecular subtypes of breast cancer. Using a cohort of 135 human breast tumors and their matched normal tissues, as well as 5 human-derived cell lines, we identified H3k4ac as a new prime target of SIRT1 in breast cancer. We also uncovered an inverse correlation between SIRT1 and the 3 epigenetic marks H3k4ac, H3k9ac and H4k16ac expression patterns. We showed that SIRT1 modulates the acetylation patterns of histones H3 and H4 in breast cancer. Moreover, SIRT1 regulates its H3 acetylated targets in a subtype-specific manner. Furthermore, SIRT1 siRNA-mediated knockdown increases histone acetylation levels at 6 breast cancer-related gene promoters: <em>AR</em>, <em>BRCA1</em>, <em>ERS1</em>, <em>ERS2</em>, <em>EZH2</em> and <em>EP300</em>. In summary, this report characterizes for the first time the epigenetic behavior of SIRT1 in human breast carcinoma. These novel findings point to a potential use of SIRT1 as an epigenetic therapeutic target in breast cancer.</p>',
'date' => '2018-07-17',
'pmid' => 'http://www.oncotarget.com/index.php?journal=oncotarget&page=article&op=view&path[]=25771&path[]=80619',
'doi' => '',
'modified' => '2018-08-09 10:47:58',
'created' => '2018-07-26 12:02:12',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 20 => array(
'id' => '3595',
'name' => 'Reciprocal signalling by Notch-Collagen V-CALCR retains muscle stem cells in their niche.',
'authors' => 'Baghdadi MB, Castel D, Machado L, Fukada SI, Birk DE, Relaix F, Tajbakhsh S, Mourikis P',
'description' => '<p>The cell microenvironment, which is critical for stem cell maintenance, contains both cellular and non-cellular components, including secreted growth factors and the extracellular matrix. Although Notch and other signalling pathways have previously been reported to regulate quiescence of stem cells, the composition and source of molecules that maintain the stem cell niche remain largely unknown. Here we show that adult muscle satellite (stem) cells in mice produce extracellular matrix collagens to maintain quiescence in a cell-autonomous manner. Using chromatin immunoprecipitation followed by sequencing, we identified NOTCH1/RBPJ-bound regulatory elements adjacent to specific collagen genes, the expression of which is deregulated in Notch-mutant mice. Moreover, we show that Collagen V (COLV) produced by satellite cells is a critical component of the quiescent niche, as depletion of COLV by conditional deletion of the Col5a1 gene leads to anomalous cell cycle entry and gradual diminution of the stem cell pool. Notably, the interaction of COLV with satellite cells is mediated by the Calcitonin receptor, for which COLV acts as a surrogate local ligand. Systemic administration of a calcitonin derivative is sufficient to rescue the quiescence and self-renewal defects found in COLV-null satellite cells. This study reveals a Notch-COLV-Calcitonin receptor signalling cascade that maintains satellite cells in a quiescent state in a cell-autonomous fashion, and raises the possibility that similar reciprocal mechanisms act in diverse stem cell populations.</p>',
'date' => '2018-05-23',
'pmid' => 'http://www.pubmed.gov/29795344',
'doi' => '10.1038/s41586-018-0144-9',
'modified' => '2019-04-17 15:12:55',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 21 => array(
'id' => '3589',
'name' => 'A new metabolic gene signature in prostate cancer regulated by JMJD3 and EZH2.',
'authors' => 'Daures M, Idrissou M, Judes G, Rifaï K, Penault-Llorca F, Bignon YJ, Guy L, Bernard-Gallon D',
'description' => '<p>Histone methylation is essential for gene expression control. Trimethylated lysine 27 of histone 3 (H3K27me3) is controlled by the balance between the activities of JMJD3 demethylase and EZH2 methyltransferase. This epigenetic mark has been shown to be deregulated in prostate cancer, and evidence shows H3K27me3 enrichment on gene promoters in prostate cancer. To study the impact of this enrichment, a transcriptomic analysis with TaqMan Low Density Array (TLDA) of several genes was studied on prostate biopsies divided into three clinical grades: normal ( = 23) and two tumor groups that differed in their aggressiveness (Gleason score ≤ 7 ( = 20) and >7 ( = 19)). ANOVA demonstrated that expression of the gene set was upregulated in tumors and correlated with Gleason score, thus discriminating between the three clinical groups. Six genes involved in key cellular processes stood out: , , , , and . Chromatin immunoprecipitation demonstrated collocation of EZH2 and JMJD3 on gene promoters that was dependent on disease stage. Gene set expression was also evaluated on prostate cancer cell lines (DU 145, PC-3 and LNCaP) treated with an inhibitor of JMJD3 (GSK-J4) or EZH2 (DZNeP) to study their involvement in gene regulation. Results showed a difference in GSK-J4 sensitivity under PTEN status of cell lines and an opposite gene expression profile according to androgen status of cells. In summary, our data describe the impacts of JMJD3 and EZH2 on a new gene signature involved in prostate cancer that may help identify diagnostic and therapeutic targets in prostate cancer.</p>',
'date' => '2018-05-04',
'pmid' => 'http://www.pubmed.gov/29805743',
'doi' => '10.18632/oncotarget.25182',
'modified' => '2019-04-17 15:21:33',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 22 => array(
'id' => '3477',
'name' => 'Contrasting epigenetic states of heterochromatin in the different types of mouse pluripotent stem cells.',
'authors' => 'Tosolini M, Brochard V, Adenot P, Chebrout M, Grillo G, Navia V, Beaujean N, Francastel C, Bonnet-Garnier A, Jouneau A',
'description' => '<p>Mouse embryonic stem cells (ESCs) and epiblast stem cells (EpiSCs) represent naive and primed pluripotency states, respectively, and are maintained in vitro by specific signalling pathways. Furthermore, ESCs cultured in serum-free medium with two kinase inhibitors (2i-ESCs) are thought to be the ground naïve pluripotent state. Here, we present a comparative study of the epigenetic and transcriptional states of pericentromeric heterochromatin satellite sequences found in these pluripotent states. We show that 2i-ESCs are distinguished from other pluripotent cells by a prominent enrichment in H3K27me3 and low levels of DNA methylation at pericentromeric heterochromatin. In contrast, serum-containing ESCs exhibit higher levels of major satellite repeat transcription, which is lower in 2i-ESCs and even more repressed in primed EpiSCs. Removal of either DNA methylation or H3K9me3 at PCH in 2i-ESCs leads to enhanced deposition of H3K27me3 with few changes in satellite transcript levels. In contrast, their removal in EpiSCs does not lead to deposition of H3K27me3 but rather removes transcriptional repression. Altogether, our data show that the epigenetic state of PCH is modified during transition from naive to primed pluripotency states towards a more repressive state, which tightly represses the transcription of satellite repeats.</p>',
'date' => '2018-04-10',
'pmid' => 'http://www.pubmed.gov/29636490',
'doi' => '10.1038/s41598-018-23822-4',
'modified' => '2019-02-15 20:26:34',
'created' => '2019-02-14 15:01:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 23 => array(
'id' => '3324',
'name' => 'Isl1 mediates mesenchymal expansion in the developing external genitalia via regulation of Bmp4, Fgf10 and Wnt5a',
'authors' => 'Ching S.T. et al.',
'description' => '<p>Genital malformations are among the most common human birth defects, and both genetic and environmental factors can contribute to these malformations. Development of the external genitalia in mammals relies on complex signaling networks, and disruption of these signaling pathways can lead to genital defects. Islet-1 (ISL1), a member of the LIM/Homeobox family of transcription factors, has been identified as a major susceptibility gene for classic bladder exstrophy in humans, a common form of the bladder exstrophy-epispadias complex (BEEC), and is implicated in a role in urinary tract development. We report that deletion of Isl1 from the genital mesenchyme in mice led to hypoplasia of the genital tubercle and prepuce, with an ectopic urethral opening and epispadias-like phenotype. These mice also developed hydroureter and hydronephrosis. Identification of ISL1 transcriptional targets via ChIP-Seq and expression analyses revealed that Isl1 regulates several important signaling pathways during embryonic genital development, including the BMP, WNT, and FGF cascades. An essential function of Isl1 during development of the external genitalia is to induce Bmp4-mediated apoptosis in the genital mesenchyme. Together, these studies demonstrate that Isl1 plays a critical role during development of the external genitalia and forms the basis for a greater understanding of the molecular mechanisms underlying the pathogenesis of BEEC and urinary tract defects in humans.</p>',
'date' => '2018-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/29126155',
'doi' => '',
'modified' => '2018-02-06 09:25:22',
'created' => '2018-02-06 09:25:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 24 => array(
'id' => '3322',
'name' => 'In Situ Fixation Redefines Quiescence and Early Activation of Skeletal Muscle Stem Cells',
'authors' => 'Machado L. et al.',
'description' => '<div class="abstract">
<h2 class="sectionTitle" tabindex="0">Summary</h2>
<div class="content">
<p>State of the art techniques have been developed to isolate and analyze cells from various tissues, aiming to capture their <em>in vivo</em> state. However, the majority of cell isolation protocols involve lengthy mechanical and enzymatic dissociation steps followed by flow cytometry, exposing cells to stress and disrupting their physiological niche. Focusing on adult skeletal muscle stem cells, we have developed a protocol that circumvents the impact of isolation procedures and captures cells in their native quiescent state. We show that current isolation protocols induce major transcriptional changes accompanied by specific histone modifications while having negligible effects on DNA methylation. In addition to proposing a protocol to avoid isolation-induced artifacts, our study reveals previously undetected quiescence and early activation genes of potential biological interest.</p>
</div>
</div>',
'date' => '2017-11-14',
'pmid' => 'http://www.cell.com/cell-reports/abstract/S2211-1247(17)31543-7',
'doi' => '',
'modified' => '2022-05-19 16:11:43',
'created' => '2018-02-02 16:36:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 25 => array(
'id' => '3263',
'name' => 'The RPMI-1640 vitamin mixture promotes bovine blastocyst development in vitro and downregulates gene expression of TXNIP with epigenetic modification of associated histones',
'authors' => 'Ikeda S.et al.',
'description' => '<p>Diverse environmental conditions surrounding preimplantation embryos, including available nutrients, affect their metabolism and development in both short- and long-term manner. Thioredoxin-interacting protein (TXNIP) is a possible marker for preimplantation stress that is implicated in in vitro fertilization- (IVF) induced long-term DOHaD effects. B vitamins, as participants in one-carbon metabolism, may affect preimplantation embryos by epigenetic alterations of metabolically and developmentally important genes. In vitro-produced bovine embryos were cultured with or without Roswell Park Memorial Institute 1640 vitamin mixture, containing B vitamins and B vitamin-like substances, from day 3 after IVF and we evaluated blastocyst development and TXNIP messenger RNA (mRNA) expression in the blastocysts by reverse transcription-quantitative polymerase chain reaction. The degree of trimethylation of histone H3 lysine 27 (H3K27me3) at TXNIP promoter was examined semi-quantitatively by chromatin immunoprecipitation polymerase chain reaction. Total H3K27me3 were also compared between the groups by Western blot analysis. The vitamin treatment significantly increased the rates of blastocyst development (P<0.05) and their hatching (P<0.001) from the zona pellucida by day 8. The mRNA expression of TXNIP was lower (P<0.01) in blastocysts in the vitamin-mixture-treated group concomitant with higher (P<0.05) level of H3K27me3 of its promoter compared with the control group. The total H3K27me3 in the vitamin-mixture-treated group was also higher (P<0.01) than that in the control group. The epigenetic control of genes related to important metabolic processes during the periconceptional period by nutritional conditions in utero and/or in vitro may have possible implication for the developmental programming during this period that may impact the welfare and production traits of farm animals.</p>',
'date' => '2017-08-02',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28764817',
'doi' => '',
'modified' => '2017-10-09 15:59:51',
'created' => '2017-10-09 15:59:51',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 26 => array(
'id' => '3193',
'name' => 'Global analysis of H3K27me3 as an epigenetic marker in prostate cancer progression',
'authors' => 'Ngollo M. et al.',
'description' => '<div class="">
<h4>BACKGROUND:</h4>
<p><abstracttext label="BACKGROUND" nlmcategory="BACKGROUND">H3K27me3 histone marks shape the inhibition of gene transcription. In prostate cancer, the deregulation of H3K27me3 marks might play a role in prostate tumor progression.</abstracttext></p>
<h4>METHODS:</h4>
<p><abstracttext label="METHODS" nlmcategory="METHODS">We investigated genome-wide H3K27me3 histone methylation profile using chromatin immunoprecipitation (ChIP) and 2X400K promoter microarrays to identify differentially-enriched regions in biopsy samples from prostate cancer patients. H3K27me3 marks were assessed in 34 prostate tumors: 11 with Gleason score > 7 (GS > 7), 10 with Gleason score ≤ 7 (GS ≤ 7), and 13 morphologically normal prostate samples.</abstracttext></p>
<h4>RESULTS:</h4>
<p><abstracttext label="RESULTS" nlmcategory="RESULTS">Here, H3K27me3 profiling identified an average of 386 enriched-genes on promoter regions in healthy control group versus 545 genes in GS ≤ 7 and 748 genes in GS > 7 group. We then ran a factorial discriminant analysis (FDA) and compared the enriched genes in prostate-tumor biopsies and normal biopsies using ANOVA to identify significantly differentially-enriched genes. The analysis identified ALG5, EXOSC8, CBX1, GRID2, GRIN3B, ING3, MYO1D, NPHP3-AS1, MSH6, FBXO11, SND1, SPATS2, TENM4 and TRA2A genes. These genes are possibly associated with prostate cancer. Notably, the H3K27me3 histone mark emerged as a novel regulatory mechanism in poor-prognosis prostate cancer.</abstracttext></p>
<h4>CONCLUSIONS:</h4>
<p><abstracttext label="CONCLUSIONS" nlmcategory="CONCLUSIONS">Our findings point to epigenetic mark H3K27me3 as an important event in prostate carcinogenesis and progression. The results reported here provide new molecular insights into the pathogenesis of prostate cancer.</abstracttext></p>
</div>',
'date' => '2017-04-12',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28403887',
'doi' => '',
'modified' => '2017-06-19 14:07:35',
'created' => '2017-06-19 14:05:03',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 27 => array(
'id' => '3156',
'name' => 'Crebbp loss cooperates with Bcl2 over-expression to promote lymphoma in mice',
'authors' => 'Idoia García-Ramírez, Saber Tadros, Inés González-Herrero, Alberto Martín-Lorenzo, Guillermo Rodríguez-Hernández, Dalia Moore, Lucía Ruiz-Roca, Oscar Blanco, Diego Alonso-López, Javier De Las Rivas, Keenan Hartert, Romain Duval, David Klinkebiel, Martin B',
'description' => '<p><span>CREBBP is targeted by inactivating mutations in follicular lymphoma (FL) and diffuse large B-cell lymphoma (DLBCL). Here, we provide evidence from transgenic mouse models that Crebbp deletion results in deficits in B-cell development and can cooperate with Bcl2 over-expression to promote B-cell lymphoma. Through transcriptional and epigenetic profiling of these B-cells we found that Crebbp inactivation was associated with broad transcriptional alterations, but no changes in the patterns of histone acetylation at the proximal regulatory regions of these genes. However, B-cells with Crebbp inactivation showed high expression of Myc and patterns of altered histone acetylation that were localized to intragenic regions, enriched for Myc DNA binding motifs, and showed Myc binding. Through the analysis of CREBBP mutations from a large cohort of primary human FL and DLBCL, we show a significant difference in the spectrum of CREBBP mutations in these two diseases, with higher frequencies of nonsense/frameshift mutations in DLBCL compared to FL. Together our data therefore provide important links between Crebbp inactivation and Bcl2 dependence, and show a role for Crebbp inactivation in the induction of Myc expression. We suggest this may parallel the role of CREBBP frameshift/nonsense mutations in DLBCL that result in loss of the protein, but may contrast the role of missense mutations in the lysine acetyltransferase domain that are more frequently observed in FL and yield an inactive protein.</span></p>',
'date' => '2017-03-05',
'pmid' => 'http://www.bloodjournal.org/content/early/2017/03/13/blood-2016-08-733469?sso-checked=true',
'doi' => 'https://doi.org/10.1182/blood-2016-08-733469',
'modified' => '2017-05-11 11:17:42',
'created' => '2017-04-10 07:56:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 28 => array(
'id' => '3151',
'name' => 'Aorta macrophage inflammatory and epigenetic changes in a murine model of obstructive sleep apnea: Potential role of CD36.',
'authors' => 'Cortese R. et al.',
'description' => '<p>Obstructive sleep apnea (OSA) affects 8-10% of the population, is characterized by chronic intermittent hypoxia (CIH), and causally associates with cardiovascular morbidities. In CIH-exposed mice, closely mimicking the chronicity of human OSA, increased accumulation and proliferation of pro-inflammatory metabolic M1-like macrophages highly expressing CD36, emerged in aorta. Transcriptomic and MeDIP-seq approaches identified activation of pro-atherogenic pathways involving a complex interplay of histone modifications in functionally-relevant biological pathways, such as inflammation and oxidative stress in aorta macrophages. Discontinuation of CIH did not elicit significant improvements in aorta wall macrophage phenotype. However, CIH-induced aorta changes were absent in CD36 knockout mice, Our results provide mechanistic insights showing that CIH exposures during sleep in absence of concurrent pro-atherogenic settings (i.e., genetic propensity or dietary manipulation) lead to the recruitment of CD36(+)<sup>high</sup> macrophages to the aortic wall and trigger atherogenesis. Furthermore, long-term CIH-induced changes may not be reversible with usual OSA treatment.</p>',
'date' => '2017-02-27',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28240319',
'doi' => '',
'modified' => '2017-03-28 09:16:02',
'created' => '2017-03-28 09:16:02',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 29 => array(
'id' => '3029',
'name' => 'Loss of Ezh2 synergizes with JAK2-V617F in initiating myeloproliferative neoplasms and promoting myelofibrosis',
'authors' => 'Shimizu T et al.',
'description' => '<p>Myeloproliferative neoplasm (MPN) patients frequently show co-occurrence of <em>JAK2</em>-V617F and mutations in epigenetic regulator genes, including <em>EZH2</em>. In this study, we show that <em>JAK2</em>-V617F and loss of <em>Ezh2</em> in hematopoietic cells contribute synergistically to the development of MPN. The MPN phenotype induced by <em>JAK2</em>-V617F was accentuated in <em>JAK2</em>-V617F<em>;Ezh2<sup>−/−</sup></em> mice, resulting in very high platelet and neutrophil counts, more advanced myelofibrosis, and reduced survival. These mice also displayed expansion of the stem cell and progenitor cell compartments and a shift of differentiation toward megakaryopoiesis at the expense of erythropoiesis. Single cell limiting dilution transplantation with bone marrow from <em>JAK2</em>-V617F<em>;Ezh2<sup>+/−</sup></em> mice showed increased reconstitution and MPN disease initiation potential compared with <em>JAK2</em>-V617F alone. RNA sequencing in <em>Ezh2</em>-deficient hematopoietic stem cells (HSCs) and megakaryocytic erythroid progenitors identified highly up-regulated genes, including <em>Lin28b</em> and <em>Hmga2</em>, and chromatin immunoprecipitation (ChIP)–quantitative PCR (qPCR) analysis of their promoters revealed decreased H3K27me3 deposition. Forced expression of <em>Hmga2</em> resulted in increased chimerism and platelet counts in recipients of retrovirally transduced HSCs. <em>JAK2</em>-V617F–expressing mice treated with an Ezh2 inhibitor showed higher platelet counts than vehicle controls. Our data support the proposed tumor suppressor function of <em>EZH2</em> in patients with MPN and call for caution when considering using Ezh2 inhibitors in MPN.</p>',
'date' => '2016-07-11',
'pmid' => 'http://jem.rupress.org/content/213/8/1479.short?rss=1',
'doi' => '',
'modified' => '2016-09-08 15:24:48',
'created' => '2016-09-08 15:24:48',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 30 => array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
[maximum depth reached]
)
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
'id' => '174',
'name' => 'MicroChIP DiaPure columns SDS GB en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-GB-en-1_0.pdf',
'countries' => 'GB',
'modified' => '2020-06-08 15:17:58',
'created' => '2020-06-08 15:17:58',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '176',
'name' => 'MicroChIP DiaPure columns SDS US en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-US-en-1_0.pdf',
'countries' => 'US',
'modified' => '2020-06-08 15:19:24',
'created' => '2020-06-08 15:19:24',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '170',
'name' => 'MicroChIP DiaPure columns SDS BE nl',
'language' => 'nl',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-nl-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:13:47',
'created' => '2020-06-08 15:13:47',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '169',
'name' => 'MicroChIP DiaPure columns SDS BE fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-fr-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:12:51',
'created' => '2020-06-08 15:12:51',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '173',
'name' => 'MicroChIP DiaPure columns SDS FR fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-FR-fr-1_0.pdf',
'countries' => 'FR',
'modified' => '2020-06-08 15:16:54',
'created' => '2020-06-08 15:16:54',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '172',
'name' => 'MicroChIP DiaPure columns SDS ES es',
'language' => 'es',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-ES-es-1_0.pdf',
'countries' => 'ES',
'modified' => '2020-06-08 15:16:02',
'created' => '2020-06-08 15:16:02',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '171',
'name' => 'MicroChIP DiaPure columns SDS DE de',
'language' => 'de',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-DE-de-1_0.pdf',
'countries' => 'DE',
'modified' => '2020-06-08 15:14:45',
'created' => '2020-06-08 15:14:45',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
)
)
)
$country = 'US'
$countries_allowed = array(
(int) 0 => 'CA',
(int) 1 => 'US',
(int) 2 => 'IE',
(int) 3 => 'GB',
(int) 4 => 'DK',
(int) 5 => 'NO',
(int) 6 => 'SE',
(int) 7 => 'FI',
(int) 8 => 'NL',
(int) 9 => 'BE',
(int) 10 => 'LU',
(int) 11 => 'FR',
(int) 12 => 'DE',
(int) 13 => 'CH',
(int) 14 => 'AT',
(int) 15 => 'ES',
(int) 16 => 'IT',
(int) 17 => 'PT'
)
$outsource = false
$other_formats = array()
$edit = ''
$testimonials = ''
$featured_testimonials = ''
$related_products = '<li>
<div class="row">
<div class="small-12 columns">
<a href="/en/p/true-microchip-kit-x16-16-rxns"><img src="/img/product/kits/chip-kit-icon.png" alt="ChIP kit icon" class="th"/></a> </div>
<div class="small-12 columns">
<div class="small-6 columns" style="padding-left:0px;padding-right:0px;margin-top:-6px;margin-left:-1px">
<span class="success label" style="">C01010132</span>
</div>
<div class="small-6 columns text-right" style="padding-left:0px;padding-right:0px;margin-top:-6px">
<!--a href="#" style="color:#B21329"><i class="fa fa-info-circle"></i></a-->
<!-- BEGIN: ADD TO CART MODAL --><div id="cartModal-1856" class="reveal-modal small" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<form action="/en/carts/add/1856" id="CartAdd/1856Form" method="post" accept-charset="utf-8"><div style="display:none;"><input type="hidden" name="_method" value="POST"/></div><input type="hidden" name="data[Cart][product_id]" value="1856" id="CartProductId"/>
<div class="row">
<div class="small-12 medium-12 large-12 columns">
<p>Add <input name="data[Cart][quantity]" placeholder="1" value="1" min="1" style="width:60px;display:inline" type="number" id="CartQuantity" required="required"/> <strong> True MicroChIP-seq Kit</strong> to my shopping cart.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="checkout" id="checkout" value="checkout" type="submit">Checkout</button> </div>
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="keepshop" id="keepshop" type="submit">Keep shopping</button> </div>
</div>
</div>
</div>
</form><a class="close-reveal-modal" aria-label="Close">×</a></div><!-- END: ADD TO CART MODAL --><a href="#" id="true-microchip-kit-x16-16-rxns" data-reveal-id="cartModal-1856" class="" style="color:#B21329"><i class="fa fa-cart-plus"></i></a>
</div>
</div>
<div class="small-12 columns" >
<h6 style="height:60px">True MicroChIP-seq Kit</h6>
</div>
</div>
</li>
'
$related = array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
'id' => '1743',
'product_id' => '1923',
'related_id' => '1856'
),
'Image' => array(
(int) 0 => array(
'id' => '1775',
'name' => 'product/kits/chip-kit-icon.png',
'alt' => 'ChIP kit icon',
'modified' => '2018-04-17 11:52:29',
'created' => '2018-03-15 15:50:34',
'ProductsImage' => array(
[maximum depth reached]
)
)
)
)
$rrbs_service = array(
(int) 0 => (int) 1894,
(int) 1 => (int) 1895
)
$chipseq_service = array(
(int) 0 => (int) 2683,
(int) 1 => (int) 1835,
(int) 2 => (int) 1836,
(int) 3 => (int) 2684,
(int) 4 => (int) 1838,
(int) 5 => (int) 1839,
(int) 6 => (int) 1856
)
$labelize = object(Closure) {
}
$old_catalog_number = ''
$country_code = 'US'
$label = '<img src="/img/banners/banner-customizer-back.png" alt=""/>'
$document = array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
'id' => '1227',
'product_id' => '1923',
'document_id' => '221'
)
)
$sds = array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
'id' => '343',
'product_id' => '1923',
'safety_sheet_id' => '175'
)
)
$publication = array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
'id' => '1173',
'product_id' => '1923',
'publication_id' => '2895'
)
)
$externalLink = ' <a href="http://www.ncbi.nlm.nih.gov/pubmed/26372092" target="_blank"><i class="fa fa-external-link"></i></a>'include - APP/View/Products/view.ctp, line 755
View::_evaluate() - CORE/Cake/View/View.php, line 971
View::_render() - CORE/Cake/View/View.php, line 933
View::render() - CORE/Cake/View/View.php, line 473
Controller::render() - CORE/Cake/Controller/Controller.php, line 963
ProductsController::slug() - APP/Controller/ProductsController.php, line 1052
ReflectionMethod::invokeArgs() - [internal], line ??
Controller::invokeAction() - CORE/Cake/Controller/Controller.php, line 491
Dispatcher::_invoke() - CORE/Cake/Routing/Dispatcher.php, line 193
Dispatcher::dispatch() - CORE/Cake/Routing/Dispatcher.php, line 167
[main] - APP/webroot/index.php, line 118
Notice (8): Undefined variable: header [APP/View/Products/view.ctp, line 755]Code Context<!-- BEGIN: REQUEST_FORM MODAL -->
<div id="request_formModal" class="reveal-modal medium" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<?= $this->element('Forms/simple_form', array('solution_of_interest' => $solution_of_interest, 'header' => $header, 'message' => $message, 'campaign_id' => $campaign_id)) ?>
$viewFile = '/home/website-server/www/app/View/Products/view.ctp'
$dataForView = array(
'language' => 'en',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'meta_title' => 'MicroChIP DiaPure columns',
'product' => array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Application' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
)
),
'Category' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Document' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
),
(int) 8 => array(
[maximum depth reached]
),
(int) 9 => array(
[maximum depth reached]
),
(int) 10 => array(
[maximum depth reached]
),
(int) 11 => array(
[maximum depth reached]
),
(int) 12 => array(
[maximum depth reached]
),
(int) 13 => array(
[maximum depth reached]
),
(int) 14 => array(
[maximum depth reached]
),
(int) 15 => array(
[maximum depth reached]
),
(int) 16 => array(
[maximum depth reached]
),
(int) 17 => array(
[maximum depth reached]
),
(int) 18 => array(
[maximum depth reached]
),
(int) 19 => array(
[maximum depth reached]
),
(int) 20 => array(
[maximum depth reached]
),
(int) 21 => array(
[maximum depth reached]
),
(int) 22 => array(
[maximum depth reached]
),
(int) 23 => array(
[maximum depth reached]
),
(int) 24 => array(
[maximum depth reached]
),
(int) 25 => array(
[maximum depth reached]
),
(int) 26 => array(
[maximum depth reached]
),
(int) 27 => array(
[maximum depth reached]
),
(int) 28 => array(
[maximum depth reached]
),
(int) 29 => array(
[maximum depth reached]
),
(int) 30 => array(
[maximum depth reached]
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
)
)
)
)
$language = 'en'
$meta_keywords = ''
$meta_description = 'MicroChIP DiaPure columns'
$meta_title = 'MicroChIP DiaPure columns'
$product = array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
[maximum depth reached]
),
'Image' => array(
[maximum depth reached]
)
)
),
'Application' => array(
(int) 0 => array(
'id' => '10',
'position' => '10',
'parent_id' => '2',
'name' => 'ChIP-qPCR',
'description' => '<div class="row">
<div class="small-12 medium-12 large-12 columns text-justify">
<p class="text-justify">Chromatin Immunoprecipitation (ChIP) coupled with quantitative PCR can be used to investigate protein-DNA interaction at known genomic binding sites. if sites are not known, qPCR primers can also be designed against potential regulatory regions such as promoters. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of performing real-time PCR is minimal. This technique is now used in a variety of life science disciplines including cellular differentiation, tumor suppressor gene silencing, and the effect of histone modifications on gene expression.</p>
<p class="text-justify"><strong>The ChIP-qPCR workflow</strong></p>
</div>
<div class="small-12 medium-12 large-12 columns text-center"><br /> <img src="https://www.diagenode.com/img/chip-qpcr-diagram.png" /></div>
<div class="small-12 medium-12 large-12 columns"><br />
<ol>
<li class="large-12 columns"><strong>Chromatin preparation: </strong>cell fixation (cross-linking) of chromatin-bound proteins such as histones or transcription factors to DNA followed by cell lysis.</li>
<li class="large-12 columns"><strong>Chromatin shearing: </strong>fragmentation of chromatin<strong> </strong>by sonication down to desired fragment size (100-500 bp)</li>
<li class="large-12 columns"><strong>Chromatin IP</strong>: protein-DNA complexe capture using<strong> <a href="https://www.diagenode.com/en/categories/chip-grade-antibodies">specific ChIP-grade antibodies</a></strong> against the histone or transcription factor of interest</li>
<li class="large-12 columns"><strong>DNA purification</strong>: chromatin reverse cross-linking and elution followed by purification<strong> </strong></li>
<li class="large-12 columns"><strong>qPCR and analysis</strong>: using previously designed primers to amplify IP'd material at specific loci</li>
</ol>
</div>
</div>
<div class="row" style="margin-top: 32px;">
<div class="small-12 medium-10 large-9 small-centered columns">
<div class="radius panel" style="background-color: #fff;">
<h3 class="text-center" style="color: #b21329;">Need guidance?</h3>
<p class="text-justify">Choose our full ChIP kits or simply choose what you need from antibodies, buffers, beads, chromatin shearing and purification reagents. With the ChIP Kit Customizer, you have complete flexibility on which components you want from our validated ChIP kits.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/which-kit-to-choose"><img src="https://www.diagenode.com/img/banners/banner-decide.png" alt="" /></a></div>
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/chip-kit-customizer-1"><img src="https://www.diagenode.com/img/banners/banner-customizer.png" alt="" /></a></div>
</div>
</div>
</div>
</div>',
'in_footer' => false,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'chip-qpcr',
'meta_keywords' => 'Chromatin immunoprecipitation,ChIP Quantitative PCR,polymerase chain reaction (PCR)',
'meta_description' => 'Diagenode's ChIP qPCR kits can be used to quantify enriched DNA after chromatin immunoprecipitation. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of',
'meta_title' => 'ChIP Quantitative PCR (ChIP-qPCR) | Diagenode',
'modified' => '2018-01-09 16:46:56',
'created' => '2014-12-11 00:22:08',
'ProductsApplication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '2',
'position' => '10',
'parent_id' => null,
'name' => 'エピジェネティクス・クロマチン解析',
'description' => '<div class="row">
<div class="large-12 columns">エピジェネティクス研究は、異なる転写パターン、遺伝子発現およびサイレンシングを引き起こすクロマチンの変化に対処します。<br /><br />クロマチンの主成分はDNA<span>およびヒストン蛋白質です。<span> </span></span>各ヒストンコア蛋白質(H2A<span>、</span>H2B<span>、</span>H3<span>および</span>H4<span>)の</span>2<span>つのコピーを</span>8<span>量体に組み込み、</span>DNA<span>で包んでヌクレオソームコアを形成させます。<span> </span></span>ヌクレオソームは、転写機械のDNA<span>への接近可能性および</span>クロマチン再構成因子を制御します。</div>
<div class="large-12 columns">
<p></p>
<p>クロマチン免疫沈降(ChIP<span>)は、関心対象の特定の蛋白質に対するゲノム結合部位の位置を解明するために使用される方法であり、遺伝子発現の制御に関する非常に貴重な洞察を提供します。<span> </span></span>ChIPは特定の抗原を含むクロマチン断片の選択的富化に関与します。 特定の蛋白質または蛋白質修飾を認識する抗体を使用して、特定の遺伝子座における抗原の相対存在量を決定します。</p>
<p>ChIP-seq<span>および</span>ChIP-qPCR<span>は、蛋白質</span>-DNA<span>結合部位の同定を可能にする技術です。</span></p>
<p> </p>
</div>
</div>',
'in_footer' => true,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'epigenetics-chromatin-study',
'meta_keywords' => 'Chromatin Immunoprecipitation Sequencing,ChIP-Seq,ChIP-seq grade antibodies,DNA purification,qPCR,Shearing of chromatin',
'meta_description' => 'Diagenodeは、ChIP-Seqおよびクロマチン免疫沈降(ChIP)専用キットと抗体を定量PCRアプリケーションと組み合わせて幅広く提供しています。',
'meta_title' => 'エピジェネティクス - クロマチン免疫沈降(ChIP)| Diagenode',
'modified' => '2018-01-11 09:58:47',
'created' => '2015-02-20 16:10:25',
'ProductsApplication' => array(
[maximum depth reached]
)
)
),
'Category' => array(
(int) 0 => array(
'id' => '61',
'position' => '2',
'parent_id' => '14',
'name' => 'Column-based DNA purification',
'description' => '',
'no_promo' => false,
'in_menu' => false,
'online' => true,
'tabular' => true,
'hide' => false,
'all_format' => false,
'is_antibody' => false,
'slug' => 'column-based-dna-purification',
'cookies_tag_id' => null,
'meta_keywords' => 'MicroChIP DiaPure columns,diagenode',
'meta_description' => 'Diagenode provides Column-based DNA purification for downstream analysis.',
'meta_title' => 'Column-based DNA purification | Diagenode',
'modified' => '2016-01-21 13:49:21',
'created' => '2015-07-08 10:31:04',
'ProductsCategory' => array(
[maximum depth reached]
),
'CookiesTag' => array([maximum depth reached])
)
),
'Document' => array(
(int) 0 => array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
[maximum depth reached]
)
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
'id' => '5033',
'name' => 'Trithorax regulates long-term memory in Drosophila through epigenetic maintenance of mushroom body metabolic state and translation capacity',
'authors' => 'Nicholas Raun et al.',
'description' => '<p><span>The role of epigenetics and chromatin in the maintenance of postmitotic neuronal cell identities is not well understood. Here, we show that the histone methyltransferase Trithorax (Trx) is required in postmitotic memory neurons of the </span><em>Drosophila</em><span><span> </span>mushroom body (MB) to enable their capacity for long-term memory (LTM), but not short-term memory (STM). Using MB-specific RNA-, ChIP-, and ATAC-sequencing, we find that Trx maintains homeostatic expression of several non-canonical MB-enriched transcripts, including the orphan nuclear receptor<span> </span></span><em>Hr51</em><span>, and the metabolic enzyme<span> </span></span><em>lactate dehydrogenase</em><span><span> </span>(</span><em>Ldh)</em><span>. Through these key targets, Trx facilitates a metabolic state characterized by high lactate levels in MBγ neurons. This metabolic state supports a high capacity for protein translation, a process that is essential for LTM, but not STM. These data suggest that Trx, a classic regulator of cell type specification during development, has additional functions in maintaining underappreciated aspects of postmitotic neuron identity, such as metabolic state. Our work supports a body of evidence suggesting that a high capacity for energy metabolism is an essential cell identity characteristic for neurons that mediate LTM.</span></p>',
'date' => '2025-01-27',
'pmid' => 'https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.3003004',
'doi' => 'https://doi.org/10.1371/journal.pbio.3003004',
'modified' => '2025-01-30 10:12:42',
'created' => '2025-01-30 10:12:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '4983',
'name' => 'Integrated multi-omics analysis of PBX1 in mouse adult neural stem- and progenitor cells identifies a transcriptional module that functionally links PBX1 to TCF3/4',
'authors' => 'Vera Laub et al.',
'description' => '<p><span>Developmental transcription factors act in networks, but how these networks achieve cell- and tissue specificity is still poorly understood. Here, we explored pre-B cell leukemia homeobox 1 (PBX1) in adult neurogenesis combining genomic, transcriptomic, and proteomic approaches. ChIP-seq analysis uncovered PBX1 binding to numerous genomic sites. Integration of PBX1 ChIP-seq with ATAC-seq data predicted interaction partners, which were subsequently validated by mass spectrometry. Whole transcriptome spatial RNA analysis revealed shared expression dynamics of </span><em>Pbx1</em><span><span> </span>and interacting factors. Among these were class I bHLH proteins TCF3 and TCF4. RNA-seq following<span> </span></span><em>Pbx1</em><span>,<span> </span></span><em>Tcf3</em><span><span> </span>or<span> </span></span><em>Tcf4</em><span><span> </span>knockdown identified proliferation- and differentiation associated genes as shared targets, while sphere formation assays following knockdown argued for functional cooperativity of PBX1 and TCF3 in progenitor cell proliferation. Notably, while physiological PBX1-TCF interaction has not yet been described, chromosomal translocation resulting in genomic<span> </span></span><em>TCF3::PBX1</em><span><span> </span>fusion characterizes a subtype of acute lymphoblastic leukemia. Introducing<span> </span></span><em>Pbx1</em><span><span> </span>into Nalm6 cells, a pre-B cell line expressing<span> </span></span><em>TCF3</em><span><span> </span>but lacking<span> </span></span><em>PBX1</em><span>, upregulated the leukemogenic genes<span> </span></span><em>BLK</em><span><span> </span>and<span> </span></span><em>NOTCH3</em><span>, arguing that functional PBX1-TCF cooperativity likely extends to hematopoiesis. Our study hence uncovers a transcriptional module orchestrating the balance between progenitor cell proliferation and differentiation in adult neurogenesis with potential implications for leukemia etiology.</span></p>',
'date' => '2024-10-08',
'pmid' => 'https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkae864/7815639',
'doi' => 'https://doi.org/10.1093/nar/gkae864',
'modified' => '2024-10-11 10:02:42',
'created' => '2024-10-11 10:02:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '4964',
'name' => 'A PRE loop at the dac locus acts as a topological chromatin structure that restricts and specifies enhancer–promoter communication',
'authors' => 'Denaud S. et al.',
'description' => '<p><span>Three-dimensional (3D) genome folding has a fundamental role in the regulation of developmental genes by facilitating or constraining chromatin interactions between </span><i>cis</i><span>-regulatory elements (CREs). Polycomb response elements (PREs) are a specific kind of CRE involved in the memory of transcriptional states in<span> </span></span><i>Drosophila melanogaster</i><span>. PREs act as nucleation sites for Polycomb group (PcG) proteins, which deposit the repressive histone mark H3K27me3, leading to the formation of a class of topologically associating domain (TAD) called a Polycomb domain. PREs can establish looping contacts that stabilize the gene repression of key developmental genes during development. However, the mechanism by which PRE loops fine-tune gene expression is unknown. Using clustered regularly interspaced short palindromic repeats and Cas9 genome engineering, we specifically perturbed PRE contacts or enhancer function and used complementary approaches including 4C-seq, Hi-C and Hi-M to analyze how chromatin architecture perturbation affects gene expression. Our results suggest that the PRE loop at the<span> </span></span><i>dac</i><span><span> </span>gene locus acts as a constitutive 3D chromatin scaffold during<span> </span></span><i>Drosophila</i><span><span> </span>development that forms independently of gene expression states and has a versatile function; it restricts enhancer–promoter communication and contributes to enhancer specificity.</span></p>',
'date' => '2024-08-16',
'pmid' => 'https://www.nature.com/articles/s41594-024-01375-7',
'doi' => 'https://doi.org/10.1038/s41594-024-01375-7',
'modified' => '2024-09-02 10:20:28',
'created' => '2024-09-02 10:20:28',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '4954',
'name' => 'A multiomic atlas of the aging hippocampus reveals molecular changes in response to environmental enrichment',
'authors' => 'Perez R. F. at al. ',
'description' => '<p><span>Aging involves the deterioration of organismal function, leading to the emergence of multiple pathologies. Environmental stimuli, including lifestyle, can influence the trajectory of this process and may be used as tools in the pursuit of healthy aging. To evaluate the role of epigenetic mechanisms in this context, we have generated bulk tissue and single cell multi-omic maps of the male mouse dorsal hippocampus in young and old animals exposed to environmental stimulation in the form of enriched environments. We present a molecular atlas of the aging process, highlighting two distinct axes, related to inflammation and to the dysregulation of mRNA metabolism, at the functional RNA and protein level. Additionally, we report the alteration of heterochromatin domains, including the loss of bivalent chromatin and the uncovering of a heterochromatin-switch phenomenon whereby constitutive heterochromatin loss is partially mitigated through gains in facultative heterochromatin. Notably, we observed the multi-omic reversal of a great number of aging-associated alterations in the context of environmental enrichment, which was particularly linked to glial and oligodendrocyte pathways. In conclusion, our work describes the epigenomic landscape of environmental stimulation in the context of aging and reveals how lifestyle intervention can lead to the multi-layered reversal of aging-associated decline.</span></p>',
'date' => '2024-07-16',
'pmid' => 'https://www.nature.com/articles/s41467-024-49608-z',
'doi' => 'https://doi.org/10.1038/s41467-024-49608-z',
'modified' => '2024-07-29 11:33:49',
'created' => '2024-07-29 11:33:49',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '4927',
'name' => 'Inflammatory stress-mediated chromatin changes underlie dysfunction in endothelial cells',
'authors' => 'Liu H. et al.',
'description' => '<p><span>Inflammatory stresses underlie endothelial dysfunction and contribute to the development of chronic cardiovascular disorders such as atherosclerosis and vascular fibrosis. The initial transcriptional response of endothelial cells to pro-inflammatory cytokines such as TNF-alpha is well established. However, very few studies uncover the effects of inflammatory stresses on chromatin architecture. We used integrative analysis of ATAC-seq and RNA-seq data to investigate chromatin alterations in human endothelial cells in response to TNF-alpha and febrile-range heat stress exposure. Multi-omics data analysis suggests a correlation between the transcription of stress-related genes and endothelial dysfunction drivers with chromatin regions exhibiting differential accessibility. Moreover, microscopy identified the dynamics in the nuclear organization, specifically, the changes in a subset of heterochromatic nucleoli-associated chromatin domains, the centromeres. Upon inflammatory stress exposure, the centromeres decreased association with nucleoli in a p38-dependent manner and increased the number of transcripts from pericentromeric regions. Overall, we provide two lines of evidence that suggest chromatin alterations in vascular endothelial cells during inflammatory stresses.</span></p>',
'date' => '2023-10-16',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10614786/',
'doi' => '10.1101/2023.10.11.561959',
'modified' => '2024-03-27 15:15:21',
'created' => '2024-03-27 15:15:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '4862',
'name' => 'Mutant FUS induces chromatin reorganization in the hippocampus andalters memory processes.',
'authors' => 'Tzeplaeff L. et al.',
'description' => '<p>Cytoplasmic mislocalization of the nuclear Fused in Sarcoma (FUS) protein is associated to amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Cytoplasmic FUS accumulation is recapitulated in the frontal cortex and spinal cord of heterozygous Fus mice. Yet, the mechanisms linking FUS mislocalization to hippocampal function and memory formation are still not characterized. Herein, we show that in these mice, the hippocampus paradoxically displays nuclear FUS accumulation. Multi-omic analyses showed that FUS binds to a set of genes characterized by the presence of an ETS/ELK-binding motifs, and involved in RNA metabolism, transcription, ribosome/mitochondria and chromatin organization. Importantly, hippocampal nuclei showed a decompaction of the neuronal chromatin at highly expressed genes and an inappropriate transcriptomic response was observed after spatial training of Fus mice. Furthermore, these mice lacked precision in a hippocampal-dependent spatial memory task and displayed decreased dendritic spine density. These studies shows that mutated FUS affects epigenetic regulation of the chromatin landscape in hippocampal neurons, which could participate in FTD/ALS pathogenic events. These data call for further investigation in the neurological phenotype of FUS-related diseases and open therapeutic strategies towards epigenetic drugs.</p>',
'date' => '2023-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/37327984',
'doi' => '10.1016/j.pneurobio.2023.102483',
'modified' => '2023-08-01 14:55:49',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '4733',
'name' => 'FXR inhibition may protect from SARS-CoV-2 infection by reducingACE2.',
'authors' => 'Brevini Teresa et al.',
'description' => '<p>Prevention of SARS-CoV-2 infection through the modulation of viral host receptors, such as ACE2, could represent a new chemoprophylactic approach for COVID-19 complementing vaccination. However, the mechanisms controlling ACE2 expression remain elusive. Here, we identify the farnesoid X receptor (FXR) as a direct regulator of ACE2 transcription in multiple COVID19-affected tissues, including the gastrointestinal and respiratory systems. We then use the over-the-counter compound z-guggulsterone (ZGG) and the off-patent drug ursodeoxycholic acid (UDCA) to reduce FXR signalling and downregulate ACE2 in human lung, cholangiocyte and intestinal organoids and in the corresponding tissues in mice and hamsters. We demonstrate that UDCA-mediated ACE2 downregulation reduces susceptibility to SARS-CoV-2 infection in vitro, in vivo and in human lungs and livers perfused ex situ. Furthermore, we illustrate that UDCA reduces ACE2 expression in the nasal epithelium in humans. Finally, we identify a correlation between UDCA treatment and positive clinical outcomes following SARS-CoV-2 infection using retrospective registry data, and confirm these findings in an independent validation cohort of liver transplant recipients. In conclusion, we identify a novel function of FXR in controlling ACE2 expression and provide evidence that modulation of this pathway could be beneficial for reducing SARS-CoV-2 infection, paving the road for future clinical trials.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36470304',
'doi' => '10.1038/s41586-022-05594-0',
'modified' => '2023-03-13 08:52:11',
'created' => '2023-02-28 12:19:11',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '4575',
'name' => 'Intranasal administration of Acinetobacter lwoffii in a murine model ofasthma induces IL-6-mediated protection associated with cecal microbiotachanges.',
'authors' => 'Alashkar A. B. et al.',
'description' => '<p>BACKGROUND: Early-life exposure to certain environmental bacteria including Acinetobacter lwoffii (AL) has been implicated in protection from chronic inflammatory diseases including asthma later in life. However, the underlying mechanisms at the immune-microbe interface remain largely unknown. METHODS: The effects of repeated intranasal AL exposure on local and systemic innate immune responses were investigated in wild-type and Il6 , Il10 , and Il17 mice exposed to ovalbumin-induced allergic airway inflammation. Those investigations were expanded by microbiome analyses. To assess for AL-associated changes in gene expression, the picture arising from animal data was supplemented by in vitro experiments of macrophage and T-cell responses, yielding expression and epigenetic data. RESULTS: The asthma preventive effect of AL was confirmed in the lung. Repeated intranasal AL administration triggered a proinflammatory immune response particularly characterized by elevated levels of IL-6, and consequently, IL-6 induced IL-10 production in CD4 T-cells. Both IL-6 and IL-10, but not IL-17, were required for asthma protection. AL had a profound impact on the gene regulatory landscape of CD4 T-cells which could be largely recapitulated by recombinant IL-6. AL administration also induced marked changes in the gastrointestinal microbiome but not in the lung microbiome. By comparing the effects on the microbiota according to mouse genotype and AL-treatment status, we have identified microbial taxa that were associated with either disease protection or activity. CONCLUSION: These experiments provide a novel mechanism of Acinetobacter lwoffii-induced asthma protection operating through IL-6-mediated epigenetic activation of IL-10 production and with associated effects on the intestinal microbiome.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36458896',
'doi' => '10.1111/all.15606',
'modified' => '2023-04-11 10:23:07',
'created' => '2023-02-21 09:59:46',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 8 => array(
'id' => '4836',
'name' => 'Caffeine intake exerts dual genome-wide effects on hippocampal metabolismand learning-dependent transcription.',
'authors' => 'Paiva I. et al.',
'description' => '<p>Caffeine is the most widely consumed psychoactive substance in the world. Strikingly, the molecular pathways engaged by its regular consumption remain unclear. We herein addressed the mechanisms associated with habitual (chronic) caffeine consumption in the mouse hippocampus using untargeted orthogonal omics techniques. Our results revealed that chronic caffeine exerts concerted pleiotropic effects in the hippocampus at the epigenomic, proteomic, and metabolomic levels. Caffeine lowered metabolism-related processes (e.g., at the level of metabolomics and gene expression) in bulk tissue, while it induced neuron-specific epigenetic changes at synaptic transmission/plasticity-related genes and increased experience-driven transcriptional activity. Altogether, these findings suggest that regular caffeine intake improves the signal-to-noise ratio during information encoding, in part through fine-tuning of metabolic genes, while boosting the salience of information processing during learning in neuronal circuits.</p>',
'date' => '2022-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35536645',
'doi' => '10.1172/JCI149371',
'modified' => '2023-08-01 13:52:29',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 9 => array(
'id' => '4220',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in Mouse Prostate Cancer Xenografts',
'authors' => 'Sanchez A. et al.',
'description' => '<p><strong class="sub-title">Background/aim:<span> </span></strong>Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo.</p>
<p><strong class="sub-title">Materials and methods:<span> </span></strong>Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR.</p>
<p><strong class="sub-title">Results:<span> </span></strong>JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression.</p>
<p><strong class="sub-title">Conclusion:<span> </span></strong>JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-05-01',
'pmid' => 'https://pubmed.ncbi.nlm.nih.gov/35430567/',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-04-21 11:54:21',
'created' => '2022-04-21 11:54:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 10 => array(
'id' => '4409',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in MouseProstate Cancer Xenografts.',
'authors' => 'Sanchez A. et al.',
'description' => '<p>BACKGROUND/AIM: Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo. MATERIALS AND METHODS: Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR. RESULTS: JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression. CONCLUSION: JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35430567',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-08-11 15:11:58',
'created' => '2022-08-11 12:14:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 11 => array(
'id' => '4233',
'name' => 'Autocrine Vitamin D-signaling switches off pro-inflammatory programsof Th1 cells',
'authors' => 'Chauss D.et al.',
'description' => '<p>The molecular mechanisms governing orderly shutdown and retraction of CD4+ T helper (Th)1 responses remain poorly understood. Here, we show that complement triggers contraction of Th1 responses by inducing intrinsic expression of the vitamin D (VitD) receptor (VDR) and the VitD-activating enzyme CYP27B1, permitting T cells to both activate and respond to VitD. VitD then initiated transition from pro-inflammatory IFN-γ + Th1 cells to suppressive IL-10+ cells. This process was primed by dynamic changes in the epigenetic landscape of CD4+ T cells, generating super-enhancers and recruiting several transcription factors, notably c-JUN, STAT3 and BACH2, which together with VDR shaped the transcriptional response to VitD. Accordingly, VitD did not induce IL-10 in cells with dysfunctional BACH2 or STAT3. Bronchoalveolar lavage fluid CD4+ T cells of COVID-19 patients were Th1-skewed and showed de-repression of genes down-regulated by VitD, either from lack of substrate (VitD deficiency) and/or abnormal regulation of this system.</p>',
'date' => '2021-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/34764490',
'doi' => '10.1038/s41590-021-01080-3',
'modified' => '2022-05-19 16:57:27',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 12 => array(
'id' => '4351',
'name' => 'Essential role of a ThPOK autoregulatory loop in the maintenance ofmature CD4 T cell identity and function.',
'authors' => 'Basu Jayati et al.',
'description' => '<p>The transcription factor ThPOK (encoded by the Zbtb7b gene) controls homeostasis and differentiation of mature helper T cells, while opposing their differentiation to CD4 intraepithelial lymphocytes (IELs) in the intestinal mucosa. Thus CD4 IEL differentiation requires ThPOK transcriptional repression via reactivation of the ThPOK transcriptional silencer element (Sil). In the present study, we describe a new autoregulatory loop whereby ThPOK binds to the Sil to maintain its own long-term expression in CD4 T cells. Disruption of this loop in vivo prevents persistent ThPOK expression, leads to genome-wide changes in chromatin accessibility and derepresses the colonic regulatory T (T) cell gene expression signature. This promotes selective differentiation of naive CD4 T cells into GITRPD-1CD25 (Triple) T cells and conversion to CD4 IELs in the gut, thereby providing dominant protection from colitis. Hence, the ThPOK autoregulatory loop represents a key mechanism to physiologically control ThPOK expression and T cell differentiation in the gut, with potential therapeutic relevance.</p>',
'date' => '2021-08-01',
'pmid' => 'https://doi.org/10.1038%2Fs41590-021-00980-8',
'doi' => '10.1038/s41590-021-00980-8',
'modified' => '2022-06-22 12:32:59',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 13 => array(
'id' => '4127',
'name' => 'The histone modification H3K4me3 is altered at the locus in Alzheimer'sdisease brain.',
'authors' => 'Smith, Adam et al.',
'description' => '<p>Several epigenome-wide association studies of DNA methylation have highlighted altered DNA methylation in the gene in Alzheimer's disease (AD) brain samples. However, no study has specifically examined histone modifications in the disease. We use chromatin immunoprecipitation-qPCR to quantify tri-methylation at histone 3 lysine 4 (H3K4me3) and 27 (H3K27me3) in the gene in entorhinal cortex from donors with high (n = 59) or low (n = 29) Alzheimer's disease pathology. We demonstrate decreased levels of H3K4me3, a marker of active gene transcription, with no change in H3K27me3, a marker of inactive genes. H3K4me3 is negatively correlated with DNA methylation in specific regions of the gene. Our study suggests that the gene shows altered epigenetic marks indicative of reduced gene activation in Alzheimer's disease.</p>',
'date' => '2021-02-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/33815817',
'doi' => '10.2144/fsoa-2020-0161',
'modified' => '2021-12-07 10:16:08',
'created' => '2021-12-06 15:53:19',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 14 => array(
'id' => '4062',
'name' => 'Digging Deeper into Breast Cancer Epigenetics: Insights from ChemicalInhibition of Histone Acetyltransferase TIP60 .',
'authors' => 'Idrissou, Mouhamed and Lebert, Andre and Boisnier, Tiphanie and Sanchez,Anna and Houfaf Khoufaf, Fatma Zohra and Penault-Llorca, Frédérique andBignon, Yves-Jean and Bernard-Gallon, Dominique',
'description' => '<p>Breast cancer is often sporadic due to several factors. Among them, the deregulation of epigenetic proteins may be involved. TIP60 or KAT5 is an acetyltransferase that regulates gene transcription through the chromatin structure. This pleiotropic protein acts in several cellular pathways by acetylating proteins. RNA and protein expressions of TIP60 were shown to decrease in some breast cancer subtypes, particularly in triple-negative breast cancer (TNBC), where a low expression of TIP60 was exhibited compared with luminal subtypes. In this study, the inhibition of the residual activity of TIP60 in breast cancer cell lines was investigated by using two chemical inhibitors, TH1834 and NU9056, first on the acetylation of the specific target, lysine 4 of histone 3 (H3K4) by immunoblotting, and second, by chromatin immunoprecipitation (ChIP)-qPCR (-quantitative Polymerase Chain Reaction). Subsequently, significant decreases or a trend toward decrease of H3K4ac in the different chromatin compartments were observed. In addition, the expression of 48 human nuclear receptors was studied with TaqMan Low-Density Array in these breast cancer cell lines treated with TIP60 inhibitors. The statistical analysis allowed us to comprehensively characterize the androgen receptor and receptors in TNBC cell lines after TH1834 or NU9056 treatment. The understanding of the residual activity of TIP60 in the evolution of breast cancer might be a major asset in the fight against this disease, and could allow TIP60 to be used as a biomarker or therapeutic target for breast cancer progression in the future.</p>',
'date' => '2020-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/32960142',
'doi' => '10.1089/omi.2020.0104',
'modified' => '2021-02-19 17:39:52',
'created' => '2021-02-18 10:21:53',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 15 => array(
'id' => '3969',
'name' => 'Delineating the early transcriptional specification of the mammalian trachea and esophagus.',
'authors' => 'Kuwahara A, Lewis AE, Coombes C, Leung FS, Percharde M, Bush JO',
'description' => '<p>The genome-scale transcriptional programs that specify the mammalian trachea and esophagus are unknown. Though NKX2-1 and SOX2 are hypothesized to be co-repressive master regulators of tracheoesophageal fates, this is untested at a whole transcriptomic scale and their downstream networks remain unidentified. By combining single-cell RNA-sequencing with bulk RNA-sequencing of mutants and NKX2-1 ChIP-sequencing in mouse embryos, we delineate the NKX2-1 transcriptional program in tracheoesophageal specification, and discover that the majority of the tracheal and esophageal transcriptome is NKX2-1 independent. To decouple the NKX2-1 transcriptional program from regulation by SOX2, we interrogate the expression of newly-identified tracheal and esophageal markers in / compound mutants. Finally, we discover that NKX2-1 binds directly to and and regulates their expression to control mesenchymal specification to cartilage and smooth muscle, coupling epithelial identity with mesenchymal specification. These findings create a new framework for understanding early tracheoesophageal fate specification at the genome-wide level.</p>',
'date' => '2020-06-09',
'pmid' => 'http://www.pubmed.gov/32515350',
'doi' => '10.7554/eLife.55526',
'modified' => '2020-08-12 09:32:02',
'created' => '2020-08-10 12:12:25',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 16 => array(
'id' => '3845',
'name' => 'Combinatorial action of NF-Y and TALE at embryonic enhancers defines distinct gene expression programs during zygotic genome activation in zebrafish.',
'authors' => 'Stanney W, Ladam F, Donaldson IJ, Parsons TJ, Maehr R, Bobola N, Sagerström CG',
'description' => '<p>Animal embryogenesis is initiated by maternal factors, but zygotic genome activation (ZGA) shifts regulatory control to the embryo during blastula stages. ZGA is thought to be mediated by maternally provided transcription factors (TFs), but few such TFs have been identified in vertebrates. Here we report that NF-Y and TALE TFs bind zebrafish genomic elements associated with developmental control genes already at ZGA. In particular, co-regulation by NF-Y and TALE is associated with broadly acting genes involved in transcriptional control, while regulation by either NF-Y or TALE defines genes in specific developmental processes, such that NF-Y controls a cilia gene expression program while TALE controls expression of hox genes. We also demonstrate that NF-Y and TALE-occupied genomic elements function as enhancers during embryogenesis. We conclude that combinatorial use of NF-Y and TALE at developmental enhancers permits the establishment of distinct gene expression programs at zebrafish ZGA.</p>',
'date' => '2019-12-17',
'pmid' => 'http://www.pubmed.gov/31862379',
'doi' => '10.1016/j.ydbio.2019.12.003',
'modified' => '2020-02-20 11:13:27',
'created' => '2020-02-13 10:02:44',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 17 => array(
'id' => '3744',
'name' => 'Whsc1 links pluripotency exit with mesendoderm specification.',
'authors' => 'Tian TV, Di Stefano B, Stik G, Vila-Casadesús M, Sardina JL, Vidal E, Dasti A, Segura-Morales C, De Andrés-Aguayo L, Gómez A, Goldmann J, Jaenisch R, Graf T',
'description' => '<p>How pluripotent stem cells differentiate into the main germ layers is a key question of developmental biology. Here, we show that the chromatin-related factor Whsc1 (also known as Nsd2 and MMSET) has a dual role in pluripotency exit and germ layer specification of embryonic stem cells. On induction of differentiation, a proportion of Whsc1-depleted embryonic stem cells remain entrapped in a pluripotent state and fail to form mesendoderm, although they are still capable of generating neuroectoderm. These functions of Whsc1 are independent of its methyltransferase activity. Whsc1 binds to enhancers of the mesendodermal regulators Gata4, T (Brachyury), Gata6 and Foxa2, together with Brd4, and activates the expression of these genes. Depleting each of these regulators also delays pluripotency exit, suggesting that they mediate the effects observed with Whsc1. Our data indicate that Whsc1 links silencing of the pluripotency regulatory network with activation of mesendoderm lineages.</p>',
'date' => '2019-07-01',
'pmid' => 'http://www.pubmed.gov/31235934',
'doi' => '10.1038/s41556-019-0342-1',
'modified' => '2019-08-06 16:35:35',
'created' => '2019-07-31 13:35:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 18 => array(
'id' => '3639',
'name' => 'Spatial confinement downsizes the inflammatory response of macrophages.',
'authors' => 'Jain N, Vogel V',
'description' => '<p>Macrophages respond to chemical/metabolic and physical stimuli, but their effects cannot be readily decoupled in vivo during pro-inflammatory activation. Here, we show that preventing macrophage spreading by spatial confinement, as imposed by micropatterning, microporous substrates or cell crowding, suppresses late lipopolysaccharide (LPS)-activated transcriptional programs (biomarkers IL-6, CXCL9, IL-1β, and iNOS) by mechanomodulating chromatin compaction and epigenetic alterations (HDAC3 levels and H3K36-dimethylation). Mechanistically, confinement reduces actin polymerization, thereby lowers the LPS-stimulated nuclear translocation of MRTF-A. This lowers the activity of the MRTF-A-SRF complex and subsequently downregulates the inflammatory response, as confirmed by chromatin immunoprecipitation coupled with quantitative PCR and RNA sequencing analysis. Confinement thus downregulates pro-inflammatory cytokine secretion and, well before any activation processes, the phagocytic potential of macrophages. Contrarily, early events, including activation of the LPS receptor TLR4, and downstream NF-κB and IRF3 signalling and hence the expression of early LPS-responsive genes were marginally affected by confinement. These findings have broad implications in the context of mechanobiology, inflammation and immunology, as well as in tissue engineering and regenerative medicine.</p>',
'date' => '2018-12-01',
'pmid' => 'http://www.pubmed.gov/30349032',
'doi' => '10.1038/s41563-018-0190-6',
'modified' => '2019-06-07 10:23:26',
'created' => '2019-06-06 12:11:18',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 19 => array(
'id' => '3379',
'name' => 'SIRT1-dependent epigenetic regulation of H3 and H4 histone acetylation in human breast cancer',
'authors' => 'Khaldoun Rifaï et al.',
'description' => '<p>Breast cancer is the most frequently diagnosed malignancy in women worldwide. It is well established that the complexity of carcinogenesis involves profound epigenetic deregulations that contribute to the tumorigenesis process. Deregulated H3 and H4 acetylated histone marks are amongst those alterations. Sirtuin-1 (SIRT1) is a class-III histone deacetylase deeply involved in apoptosis, genomic stability, gene expression regulation and breast tumorigenesis. However, the underlying molecular mechanism by which SIRT1 regulates H3 and H4 acetylated marks, and consequently cancer-related gene expression in breast cancer, remains uncharacterized. In this study, we elucidated SIRT1 epigenetic role and analyzed the link between the latter and histones H3 and H4 epigenetic marks in all 5 molecular subtypes of breast cancer. Using a cohort of 135 human breast tumors and their matched normal tissues, as well as 5 human-derived cell lines, we identified H3k4ac as a new prime target of SIRT1 in breast cancer. We also uncovered an inverse correlation between SIRT1 and the 3 epigenetic marks H3k4ac, H3k9ac and H4k16ac expression patterns. We showed that SIRT1 modulates the acetylation patterns of histones H3 and H4 in breast cancer. Moreover, SIRT1 regulates its H3 acetylated targets in a subtype-specific manner. Furthermore, SIRT1 siRNA-mediated knockdown increases histone acetylation levels at 6 breast cancer-related gene promoters: <em>AR</em>, <em>BRCA1</em>, <em>ERS1</em>, <em>ERS2</em>, <em>EZH2</em> and <em>EP300</em>. In summary, this report characterizes for the first time the epigenetic behavior of SIRT1 in human breast carcinoma. These novel findings point to a potential use of SIRT1 as an epigenetic therapeutic target in breast cancer.</p>',
'date' => '2018-07-17',
'pmid' => 'http://www.oncotarget.com/index.php?journal=oncotarget&page=article&op=view&path[]=25771&path[]=80619',
'doi' => '',
'modified' => '2018-08-09 10:47:58',
'created' => '2018-07-26 12:02:12',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 20 => array(
'id' => '3595',
'name' => 'Reciprocal signalling by Notch-Collagen V-CALCR retains muscle stem cells in their niche.',
'authors' => 'Baghdadi MB, Castel D, Machado L, Fukada SI, Birk DE, Relaix F, Tajbakhsh S, Mourikis P',
'description' => '<p>The cell microenvironment, which is critical for stem cell maintenance, contains both cellular and non-cellular components, including secreted growth factors and the extracellular matrix. Although Notch and other signalling pathways have previously been reported to regulate quiescence of stem cells, the composition and source of molecules that maintain the stem cell niche remain largely unknown. Here we show that adult muscle satellite (stem) cells in mice produce extracellular matrix collagens to maintain quiescence in a cell-autonomous manner. Using chromatin immunoprecipitation followed by sequencing, we identified NOTCH1/RBPJ-bound regulatory elements adjacent to specific collagen genes, the expression of which is deregulated in Notch-mutant mice. Moreover, we show that Collagen V (COLV) produced by satellite cells is a critical component of the quiescent niche, as depletion of COLV by conditional deletion of the Col5a1 gene leads to anomalous cell cycle entry and gradual diminution of the stem cell pool. Notably, the interaction of COLV with satellite cells is mediated by the Calcitonin receptor, for which COLV acts as a surrogate local ligand. Systemic administration of a calcitonin derivative is sufficient to rescue the quiescence and self-renewal defects found in COLV-null satellite cells. This study reveals a Notch-COLV-Calcitonin receptor signalling cascade that maintains satellite cells in a quiescent state in a cell-autonomous fashion, and raises the possibility that similar reciprocal mechanisms act in diverse stem cell populations.</p>',
'date' => '2018-05-23',
'pmid' => 'http://www.pubmed.gov/29795344',
'doi' => '10.1038/s41586-018-0144-9',
'modified' => '2019-04-17 15:12:55',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 21 => array(
'id' => '3589',
'name' => 'A new metabolic gene signature in prostate cancer regulated by JMJD3 and EZH2.',
'authors' => 'Daures M, Idrissou M, Judes G, Rifaï K, Penault-Llorca F, Bignon YJ, Guy L, Bernard-Gallon D',
'description' => '<p>Histone methylation is essential for gene expression control. Trimethylated lysine 27 of histone 3 (H3K27me3) is controlled by the balance between the activities of JMJD3 demethylase and EZH2 methyltransferase. This epigenetic mark has been shown to be deregulated in prostate cancer, and evidence shows H3K27me3 enrichment on gene promoters in prostate cancer. To study the impact of this enrichment, a transcriptomic analysis with TaqMan Low Density Array (TLDA) of several genes was studied on prostate biopsies divided into three clinical grades: normal ( = 23) and two tumor groups that differed in their aggressiveness (Gleason score ≤ 7 ( = 20) and >7 ( = 19)). ANOVA demonstrated that expression of the gene set was upregulated in tumors and correlated with Gleason score, thus discriminating between the three clinical groups. Six genes involved in key cellular processes stood out: , , , , and . Chromatin immunoprecipitation demonstrated collocation of EZH2 and JMJD3 on gene promoters that was dependent on disease stage. Gene set expression was also evaluated on prostate cancer cell lines (DU 145, PC-3 and LNCaP) treated with an inhibitor of JMJD3 (GSK-J4) or EZH2 (DZNeP) to study their involvement in gene regulation. Results showed a difference in GSK-J4 sensitivity under PTEN status of cell lines and an opposite gene expression profile according to androgen status of cells. In summary, our data describe the impacts of JMJD3 and EZH2 on a new gene signature involved in prostate cancer that may help identify diagnostic and therapeutic targets in prostate cancer.</p>',
'date' => '2018-05-04',
'pmid' => 'http://www.pubmed.gov/29805743',
'doi' => '10.18632/oncotarget.25182',
'modified' => '2019-04-17 15:21:33',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 22 => array(
'id' => '3477',
'name' => 'Contrasting epigenetic states of heterochromatin in the different types of mouse pluripotent stem cells.',
'authors' => 'Tosolini M, Brochard V, Adenot P, Chebrout M, Grillo G, Navia V, Beaujean N, Francastel C, Bonnet-Garnier A, Jouneau A',
'description' => '<p>Mouse embryonic stem cells (ESCs) and epiblast stem cells (EpiSCs) represent naive and primed pluripotency states, respectively, and are maintained in vitro by specific signalling pathways. Furthermore, ESCs cultured in serum-free medium with two kinase inhibitors (2i-ESCs) are thought to be the ground naïve pluripotent state. Here, we present a comparative study of the epigenetic and transcriptional states of pericentromeric heterochromatin satellite sequences found in these pluripotent states. We show that 2i-ESCs are distinguished from other pluripotent cells by a prominent enrichment in H3K27me3 and low levels of DNA methylation at pericentromeric heterochromatin. In contrast, serum-containing ESCs exhibit higher levels of major satellite repeat transcription, which is lower in 2i-ESCs and even more repressed in primed EpiSCs. Removal of either DNA methylation or H3K9me3 at PCH in 2i-ESCs leads to enhanced deposition of H3K27me3 with few changes in satellite transcript levels. In contrast, their removal in EpiSCs does not lead to deposition of H3K27me3 but rather removes transcriptional repression. Altogether, our data show that the epigenetic state of PCH is modified during transition from naive to primed pluripotency states towards a more repressive state, which tightly represses the transcription of satellite repeats.</p>',
'date' => '2018-04-10',
'pmid' => 'http://www.pubmed.gov/29636490',
'doi' => '10.1038/s41598-018-23822-4',
'modified' => '2019-02-15 20:26:34',
'created' => '2019-02-14 15:01:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 23 => array(
'id' => '3324',
'name' => 'Isl1 mediates mesenchymal expansion in the developing external genitalia via regulation of Bmp4, Fgf10 and Wnt5a',
'authors' => 'Ching S.T. et al.',
'description' => '<p>Genital malformations are among the most common human birth defects, and both genetic and environmental factors can contribute to these malformations. Development of the external genitalia in mammals relies on complex signaling networks, and disruption of these signaling pathways can lead to genital defects. Islet-1 (ISL1), a member of the LIM/Homeobox family of transcription factors, has been identified as a major susceptibility gene for classic bladder exstrophy in humans, a common form of the bladder exstrophy-epispadias complex (BEEC), and is implicated in a role in urinary tract development. We report that deletion of Isl1 from the genital mesenchyme in mice led to hypoplasia of the genital tubercle and prepuce, with an ectopic urethral opening and epispadias-like phenotype. These mice also developed hydroureter and hydronephrosis. Identification of ISL1 transcriptional targets via ChIP-Seq and expression analyses revealed that Isl1 regulates several important signaling pathways during embryonic genital development, including the BMP, WNT, and FGF cascades. An essential function of Isl1 during development of the external genitalia is to induce Bmp4-mediated apoptosis in the genital mesenchyme. Together, these studies demonstrate that Isl1 plays a critical role during development of the external genitalia and forms the basis for a greater understanding of the molecular mechanisms underlying the pathogenesis of BEEC and urinary tract defects in humans.</p>',
'date' => '2018-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/29126155',
'doi' => '',
'modified' => '2018-02-06 09:25:22',
'created' => '2018-02-06 09:25:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 24 => array(
'id' => '3322',
'name' => 'In Situ Fixation Redefines Quiescence and Early Activation of Skeletal Muscle Stem Cells',
'authors' => 'Machado L. et al.',
'description' => '<div class="abstract">
<h2 class="sectionTitle" tabindex="0">Summary</h2>
<div class="content">
<p>State of the art techniques have been developed to isolate and analyze cells from various tissues, aiming to capture their <em>in vivo</em> state. However, the majority of cell isolation protocols involve lengthy mechanical and enzymatic dissociation steps followed by flow cytometry, exposing cells to stress and disrupting their physiological niche. Focusing on adult skeletal muscle stem cells, we have developed a protocol that circumvents the impact of isolation procedures and captures cells in their native quiescent state. We show that current isolation protocols induce major transcriptional changes accompanied by specific histone modifications while having negligible effects on DNA methylation. In addition to proposing a protocol to avoid isolation-induced artifacts, our study reveals previously undetected quiescence and early activation genes of potential biological interest.</p>
</div>
</div>',
'date' => '2017-11-14',
'pmid' => 'http://www.cell.com/cell-reports/abstract/S2211-1247(17)31543-7',
'doi' => '',
'modified' => '2022-05-19 16:11:43',
'created' => '2018-02-02 16:36:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 25 => array(
'id' => '3263',
'name' => 'The RPMI-1640 vitamin mixture promotes bovine blastocyst development in vitro and downregulates gene expression of TXNIP with epigenetic modification of associated histones',
'authors' => 'Ikeda S.et al.',
'description' => '<p>Diverse environmental conditions surrounding preimplantation embryos, including available nutrients, affect their metabolism and development in both short- and long-term manner. Thioredoxin-interacting protein (TXNIP) is a possible marker for preimplantation stress that is implicated in in vitro fertilization- (IVF) induced long-term DOHaD effects. B vitamins, as participants in one-carbon metabolism, may affect preimplantation embryos by epigenetic alterations of metabolically and developmentally important genes. In vitro-produced bovine embryos were cultured with or without Roswell Park Memorial Institute 1640 vitamin mixture, containing B vitamins and B vitamin-like substances, from day 3 after IVF and we evaluated blastocyst development and TXNIP messenger RNA (mRNA) expression in the blastocysts by reverse transcription-quantitative polymerase chain reaction. The degree of trimethylation of histone H3 lysine 27 (H3K27me3) at TXNIP promoter was examined semi-quantitatively by chromatin immunoprecipitation polymerase chain reaction. Total H3K27me3 were also compared between the groups by Western blot analysis. The vitamin treatment significantly increased the rates of blastocyst development (P<0.05) and their hatching (P<0.001) from the zona pellucida by day 8. The mRNA expression of TXNIP was lower (P<0.01) in blastocysts in the vitamin-mixture-treated group concomitant with higher (P<0.05) level of H3K27me3 of its promoter compared with the control group. The total H3K27me3 in the vitamin-mixture-treated group was also higher (P<0.01) than that in the control group. The epigenetic control of genes related to important metabolic processes during the periconceptional period by nutritional conditions in utero and/or in vitro may have possible implication for the developmental programming during this period that may impact the welfare and production traits of farm animals.</p>',
'date' => '2017-08-02',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28764817',
'doi' => '',
'modified' => '2017-10-09 15:59:51',
'created' => '2017-10-09 15:59:51',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 26 => array(
'id' => '3193',
'name' => 'Global analysis of H3K27me3 as an epigenetic marker in prostate cancer progression',
'authors' => 'Ngollo M. et al.',
'description' => '<div class="">
<h4>BACKGROUND:</h4>
<p><abstracttext label="BACKGROUND" nlmcategory="BACKGROUND">H3K27me3 histone marks shape the inhibition of gene transcription. In prostate cancer, the deregulation of H3K27me3 marks might play a role in prostate tumor progression.</abstracttext></p>
<h4>METHODS:</h4>
<p><abstracttext label="METHODS" nlmcategory="METHODS">We investigated genome-wide H3K27me3 histone methylation profile using chromatin immunoprecipitation (ChIP) and 2X400K promoter microarrays to identify differentially-enriched regions in biopsy samples from prostate cancer patients. H3K27me3 marks were assessed in 34 prostate tumors: 11 with Gleason score > 7 (GS > 7), 10 with Gleason score ≤ 7 (GS ≤ 7), and 13 morphologically normal prostate samples.</abstracttext></p>
<h4>RESULTS:</h4>
<p><abstracttext label="RESULTS" nlmcategory="RESULTS">Here, H3K27me3 profiling identified an average of 386 enriched-genes on promoter regions in healthy control group versus 545 genes in GS ≤ 7 and 748 genes in GS > 7 group. We then ran a factorial discriminant analysis (FDA) and compared the enriched genes in prostate-tumor biopsies and normal biopsies using ANOVA to identify significantly differentially-enriched genes. The analysis identified ALG5, EXOSC8, CBX1, GRID2, GRIN3B, ING3, MYO1D, NPHP3-AS1, MSH6, FBXO11, SND1, SPATS2, TENM4 and TRA2A genes. These genes are possibly associated with prostate cancer. Notably, the H3K27me3 histone mark emerged as a novel regulatory mechanism in poor-prognosis prostate cancer.</abstracttext></p>
<h4>CONCLUSIONS:</h4>
<p><abstracttext label="CONCLUSIONS" nlmcategory="CONCLUSIONS">Our findings point to epigenetic mark H3K27me3 as an important event in prostate carcinogenesis and progression. The results reported here provide new molecular insights into the pathogenesis of prostate cancer.</abstracttext></p>
</div>',
'date' => '2017-04-12',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28403887',
'doi' => '',
'modified' => '2017-06-19 14:07:35',
'created' => '2017-06-19 14:05:03',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 27 => array(
'id' => '3156',
'name' => 'Crebbp loss cooperates with Bcl2 over-expression to promote lymphoma in mice',
'authors' => 'Idoia García-Ramírez, Saber Tadros, Inés González-Herrero, Alberto Martín-Lorenzo, Guillermo Rodríguez-Hernández, Dalia Moore, Lucía Ruiz-Roca, Oscar Blanco, Diego Alonso-López, Javier De Las Rivas, Keenan Hartert, Romain Duval, David Klinkebiel, Martin B',
'description' => '<p><span>CREBBP is targeted by inactivating mutations in follicular lymphoma (FL) and diffuse large B-cell lymphoma (DLBCL). Here, we provide evidence from transgenic mouse models that Crebbp deletion results in deficits in B-cell development and can cooperate with Bcl2 over-expression to promote B-cell lymphoma. Through transcriptional and epigenetic profiling of these B-cells we found that Crebbp inactivation was associated with broad transcriptional alterations, but no changes in the patterns of histone acetylation at the proximal regulatory regions of these genes. However, B-cells with Crebbp inactivation showed high expression of Myc and patterns of altered histone acetylation that were localized to intragenic regions, enriched for Myc DNA binding motifs, and showed Myc binding. Through the analysis of CREBBP mutations from a large cohort of primary human FL and DLBCL, we show a significant difference in the spectrum of CREBBP mutations in these two diseases, with higher frequencies of nonsense/frameshift mutations in DLBCL compared to FL. Together our data therefore provide important links between Crebbp inactivation and Bcl2 dependence, and show a role for Crebbp inactivation in the induction of Myc expression. We suggest this may parallel the role of CREBBP frameshift/nonsense mutations in DLBCL that result in loss of the protein, but may contrast the role of missense mutations in the lysine acetyltransferase domain that are more frequently observed in FL and yield an inactive protein.</span></p>',
'date' => '2017-03-05',
'pmid' => 'http://www.bloodjournal.org/content/early/2017/03/13/blood-2016-08-733469?sso-checked=true',
'doi' => 'https://doi.org/10.1182/blood-2016-08-733469',
'modified' => '2017-05-11 11:17:42',
'created' => '2017-04-10 07:56:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 28 => array(
'id' => '3151',
'name' => 'Aorta macrophage inflammatory and epigenetic changes in a murine model of obstructive sleep apnea: Potential role of CD36.',
'authors' => 'Cortese R. et al.',
'description' => '<p>Obstructive sleep apnea (OSA) affects 8-10% of the population, is characterized by chronic intermittent hypoxia (CIH), and causally associates with cardiovascular morbidities. In CIH-exposed mice, closely mimicking the chronicity of human OSA, increased accumulation and proliferation of pro-inflammatory metabolic M1-like macrophages highly expressing CD36, emerged in aorta. Transcriptomic and MeDIP-seq approaches identified activation of pro-atherogenic pathways involving a complex interplay of histone modifications in functionally-relevant biological pathways, such as inflammation and oxidative stress in aorta macrophages. Discontinuation of CIH did not elicit significant improvements in aorta wall macrophage phenotype. However, CIH-induced aorta changes were absent in CD36 knockout mice, Our results provide mechanistic insights showing that CIH exposures during sleep in absence of concurrent pro-atherogenic settings (i.e., genetic propensity or dietary manipulation) lead to the recruitment of CD36(+)<sup>high</sup> macrophages to the aortic wall and trigger atherogenesis. Furthermore, long-term CIH-induced changes may not be reversible with usual OSA treatment.</p>',
'date' => '2017-02-27',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28240319',
'doi' => '',
'modified' => '2017-03-28 09:16:02',
'created' => '2017-03-28 09:16:02',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 29 => array(
'id' => '3029',
'name' => 'Loss of Ezh2 synergizes with JAK2-V617F in initiating myeloproliferative neoplasms and promoting myelofibrosis',
'authors' => 'Shimizu T et al.',
'description' => '<p>Myeloproliferative neoplasm (MPN) patients frequently show co-occurrence of <em>JAK2</em>-V617F and mutations in epigenetic regulator genes, including <em>EZH2</em>. In this study, we show that <em>JAK2</em>-V617F and loss of <em>Ezh2</em> in hematopoietic cells contribute synergistically to the development of MPN. The MPN phenotype induced by <em>JAK2</em>-V617F was accentuated in <em>JAK2</em>-V617F<em>;Ezh2<sup>−/−</sup></em> mice, resulting in very high platelet and neutrophil counts, more advanced myelofibrosis, and reduced survival. These mice also displayed expansion of the stem cell and progenitor cell compartments and a shift of differentiation toward megakaryopoiesis at the expense of erythropoiesis. Single cell limiting dilution transplantation with bone marrow from <em>JAK2</em>-V617F<em>;Ezh2<sup>+/−</sup></em> mice showed increased reconstitution and MPN disease initiation potential compared with <em>JAK2</em>-V617F alone. RNA sequencing in <em>Ezh2</em>-deficient hematopoietic stem cells (HSCs) and megakaryocytic erythroid progenitors identified highly up-regulated genes, including <em>Lin28b</em> and <em>Hmga2</em>, and chromatin immunoprecipitation (ChIP)–quantitative PCR (qPCR) analysis of their promoters revealed decreased H3K27me3 deposition. Forced expression of <em>Hmga2</em> resulted in increased chimerism and platelet counts in recipients of retrovirally transduced HSCs. <em>JAK2</em>-V617F–expressing mice treated with an Ezh2 inhibitor showed higher platelet counts than vehicle controls. Our data support the proposed tumor suppressor function of <em>EZH2</em> in patients with MPN and call for caution when considering using Ezh2 inhibitors in MPN.</p>',
'date' => '2016-07-11',
'pmid' => 'http://jem.rupress.org/content/213/8/1479.short?rss=1',
'doi' => '',
'modified' => '2016-09-08 15:24:48',
'created' => '2016-09-08 15:24:48',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 30 => array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
[maximum depth reached]
)
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
'id' => '174',
'name' => 'MicroChIP DiaPure columns SDS GB en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-GB-en-1_0.pdf',
'countries' => 'GB',
'modified' => '2020-06-08 15:17:58',
'created' => '2020-06-08 15:17:58',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '176',
'name' => 'MicroChIP DiaPure columns SDS US en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-US-en-1_0.pdf',
'countries' => 'US',
'modified' => '2020-06-08 15:19:24',
'created' => '2020-06-08 15:19:24',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '170',
'name' => 'MicroChIP DiaPure columns SDS BE nl',
'language' => 'nl',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-nl-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:13:47',
'created' => '2020-06-08 15:13:47',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '169',
'name' => 'MicroChIP DiaPure columns SDS BE fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-fr-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:12:51',
'created' => '2020-06-08 15:12:51',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '173',
'name' => 'MicroChIP DiaPure columns SDS FR fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-FR-fr-1_0.pdf',
'countries' => 'FR',
'modified' => '2020-06-08 15:16:54',
'created' => '2020-06-08 15:16:54',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '172',
'name' => 'MicroChIP DiaPure columns SDS ES es',
'language' => 'es',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-ES-es-1_0.pdf',
'countries' => 'ES',
'modified' => '2020-06-08 15:16:02',
'created' => '2020-06-08 15:16:02',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '171',
'name' => 'MicroChIP DiaPure columns SDS DE de',
'language' => 'de',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-DE-de-1_0.pdf',
'countries' => 'DE',
'modified' => '2020-06-08 15:14:45',
'created' => '2020-06-08 15:14:45',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
)
)
)
$country = 'US'
$countries_allowed = array(
(int) 0 => 'CA',
(int) 1 => 'US',
(int) 2 => 'IE',
(int) 3 => 'GB',
(int) 4 => 'DK',
(int) 5 => 'NO',
(int) 6 => 'SE',
(int) 7 => 'FI',
(int) 8 => 'NL',
(int) 9 => 'BE',
(int) 10 => 'LU',
(int) 11 => 'FR',
(int) 12 => 'DE',
(int) 13 => 'CH',
(int) 14 => 'AT',
(int) 15 => 'ES',
(int) 16 => 'IT',
(int) 17 => 'PT'
)
$outsource = false
$other_formats = array()
$edit = ''
$testimonials = ''
$featured_testimonials = ''
$related_products = '<li>
<div class="row">
<div class="small-12 columns">
<a href="/en/p/true-microchip-kit-x16-16-rxns"><img src="/img/product/kits/chip-kit-icon.png" alt="ChIP kit icon" class="th"/></a> </div>
<div class="small-12 columns">
<div class="small-6 columns" style="padding-left:0px;padding-right:0px;margin-top:-6px;margin-left:-1px">
<span class="success label" style="">C01010132</span>
</div>
<div class="small-6 columns text-right" style="padding-left:0px;padding-right:0px;margin-top:-6px">
<!--a href="#" style="color:#B21329"><i class="fa fa-info-circle"></i></a-->
<!-- BEGIN: ADD TO CART MODAL --><div id="cartModal-1856" class="reveal-modal small" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<form action="/en/carts/add/1856" id="CartAdd/1856Form" method="post" accept-charset="utf-8"><div style="display:none;"><input type="hidden" name="_method" value="POST"/></div><input type="hidden" name="data[Cart][product_id]" value="1856" id="CartProductId"/>
<div class="row">
<div class="small-12 medium-12 large-12 columns">
<p>Add <input name="data[Cart][quantity]" placeholder="1" value="1" min="1" style="width:60px;display:inline" type="number" id="CartQuantity" required="required"/> <strong> True MicroChIP-seq Kit</strong> to my shopping cart.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="checkout" id="checkout" value="checkout" type="submit">Checkout</button> </div>
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="keepshop" id="keepshop" type="submit">Keep shopping</button> </div>
</div>
</div>
</div>
</form><a class="close-reveal-modal" aria-label="Close">×</a></div><!-- END: ADD TO CART MODAL --><a href="#" id="true-microchip-kit-x16-16-rxns" data-reveal-id="cartModal-1856" class="" style="color:#B21329"><i class="fa fa-cart-plus"></i></a>
</div>
</div>
<div class="small-12 columns" >
<h6 style="height:60px">True MicroChIP-seq Kit</h6>
</div>
</div>
</li>
'
$related = array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
'id' => '1743',
'product_id' => '1923',
'related_id' => '1856'
),
'Image' => array(
(int) 0 => array(
'id' => '1775',
'name' => 'product/kits/chip-kit-icon.png',
'alt' => 'ChIP kit icon',
'modified' => '2018-04-17 11:52:29',
'created' => '2018-03-15 15:50:34',
'ProductsImage' => array(
[maximum depth reached]
)
)
)
)
$rrbs_service = array(
(int) 0 => (int) 1894,
(int) 1 => (int) 1895
)
$chipseq_service = array(
(int) 0 => (int) 2683,
(int) 1 => (int) 1835,
(int) 2 => (int) 1836,
(int) 3 => (int) 2684,
(int) 4 => (int) 1838,
(int) 5 => (int) 1839,
(int) 6 => (int) 1856
)
$labelize = object(Closure) {
}
$old_catalog_number = ''
$country_code = 'US'
$label = '<img src="/img/banners/banner-customizer-back.png" alt=""/>'
$document = array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
'id' => '1227',
'product_id' => '1923',
'document_id' => '221'
)
)
$sds = array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
'id' => '343',
'product_id' => '1923',
'safety_sheet_id' => '175'
)
)
$publication = array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
'id' => '1173',
'product_id' => '1923',
'publication_id' => '2895'
)
)
$externalLink = ' <a href="http://www.ncbi.nlm.nih.gov/pubmed/26372092" target="_blank"><i class="fa fa-external-link"></i></a>'include - APP/View/Products/view.ctp, line 755
View::_evaluate() - CORE/Cake/View/View.php, line 971
View::_render() - CORE/Cake/View/View.php, line 933
View::render() - CORE/Cake/View/View.php, line 473
Controller::render() - CORE/Cake/Controller/Controller.php, line 963
ProductsController::slug() - APP/Controller/ProductsController.php, line 1052
ReflectionMethod::invokeArgs() - [internal], line ??
Controller::invokeAction() - CORE/Cake/Controller/Controller.php, line 491
Dispatcher::_invoke() - CORE/Cake/Routing/Dispatcher.php, line 193
Dispatcher::dispatch() - CORE/Cake/Routing/Dispatcher.php, line 167
[main] - APP/webroot/index.php, line 118
Notice (8): Undefined variable: message [APP/View/Products/view.ctp, line 755]Code Context<!-- BEGIN: REQUEST_FORM MODAL -->
<div id="request_formModal" class="reveal-modal medium" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<?= $this->element('Forms/simple_form', array('solution_of_interest' => $solution_of_interest, 'header' => $header, 'message' => $message, 'campaign_id' => $campaign_id)) ?>
$viewFile = '/home/website-server/www/app/View/Products/view.ctp'
$dataForView = array(
'language' => 'en',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'meta_title' => 'MicroChIP DiaPure columns',
'product' => array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Application' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
)
),
'Category' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Document' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
),
(int) 8 => array(
[maximum depth reached]
),
(int) 9 => array(
[maximum depth reached]
),
(int) 10 => array(
[maximum depth reached]
),
(int) 11 => array(
[maximum depth reached]
),
(int) 12 => array(
[maximum depth reached]
),
(int) 13 => array(
[maximum depth reached]
),
(int) 14 => array(
[maximum depth reached]
),
(int) 15 => array(
[maximum depth reached]
),
(int) 16 => array(
[maximum depth reached]
),
(int) 17 => array(
[maximum depth reached]
),
(int) 18 => array(
[maximum depth reached]
),
(int) 19 => array(
[maximum depth reached]
),
(int) 20 => array(
[maximum depth reached]
),
(int) 21 => array(
[maximum depth reached]
),
(int) 22 => array(
[maximum depth reached]
),
(int) 23 => array(
[maximum depth reached]
),
(int) 24 => array(
[maximum depth reached]
),
(int) 25 => array(
[maximum depth reached]
),
(int) 26 => array(
[maximum depth reached]
),
(int) 27 => array(
[maximum depth reached]
),
(int) 28 => array(
[maximum depth reached]
),
(int) 29 => array(
[maximum depth reached]
),
(int) 30 => array(
[maximum depth reached]
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
)
)
)
)
$language = 'en'
$meta_keywords = ''
$meta_description = 'MicroChIP DiaPure columns'
$meta_title = 'MicroChIP DiaPure columns'
$product = array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
[maximum depth reached]
),
'Image' => array(
[maximum depth reached]
)
)
),
'Application' => array(
(int) 0 => array(
'id' => '10',
'position' => '10',
'parent_id' => '2',
'name' => 'ChIP-qPCR',
'description' => '<div class="row">
<div class="small-12 medium-12 large-12 columns text-justify">
<p class="text-justify">Chromatin Immunoprecipitation (ChIP) coupled with quantitative PCR can be used to investigate protein-DNA interaction at known genomic binding sites. if sites are not known, qPCR primers can also be designed against potential regulatory regions such as promoters. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of performing real-time PCR is minimal. This technique is now used in a variety of life science disciplines including cellular differentiation, tumor suppressor gene silencing, and the effect of histone modifications on gene expression.</p>
<p class="text-justify"><strong>The ChIP-qPCR workflow</strong></p>
</div>
<div class="small-12 medium-12 large-12 columns text-center"><br /> <img src="https://www.diagenode.com/img/chip-qpcr-diagram.png" /></div>
<div class="small-12 medium-12 large-12 columns"><br />
<ol>
<li class="large-12 columns"><strong>Chromatin preparation: </strong>cell fixation (cross-linking) of chromatin-bound proteins such as histones or transcription factors to DNA followed by cell lysis.</li>
<li class="large-12 columns"><strong>Chromatin shearing: </strong>fragmentation of chromatin<strong> </strong>by sonication down to desired fragment size (100-500 bp)</li>
<li class="large-12 columns"><strong>Chromatin IP</strong>: protein-DNA complexe capture using<strong> <a href="https://www.diagenode.com/en/categories/chip-grade-antibodies">specific ChIP-grade antibodies</a></strong> against the histone or transcription factor of interest</li>
<li class="large-12 columns"><strong>DNA purification</strong>: chromatin reverse cross-linking and elution followed by purification<strong> </strong></li>
<li class="large-12 columns"><strong>qPCR and analysis</strong>: using previously designed primers to amplify IP'd material at specific loci</li>
</ol>
</div>
</div>
<div class="row" style="margin-top: 32px;">
<div class="small-12 medium-10 large-9 small-centered columns">
<div class="radius panel" style="background-color: #fff;">
<h3 class="text-center" style="color: #b21329;">Need guidance?</h3>
<p class="text-justify">Choose our full ChIP kits or simply choose what you need from antibodies, buffers, beads, chromatin shearing and purification reagents. With the ChIP Kit Customizer, you have complete flexibility on which components you want from our validated ChIP kits.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/which-kit-to-choose"><img src="https://www.diagenode.com/img/banners/banner-decide.png" alt="" /></a></div>
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/chip-kit-customizer-1"><img src="https://www.diagenode.com/img/banners/banner-customizer.png" alt="" /></a></div>
</div>
</div>
</div>
</div>',
'in_footer' => false,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'chip-qpcr',
'meta_keywords' => 'Chromatin immunoprecipitation,ChIP Quantitative PCR,polymerase chain reaction (PCR)',
'meta_description' => 'Diagenode's ChIP qPCR kits can be used to quantify enriched DNA after chromatin immunoprecipitation. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of',
'meta_title' => 'ChIP Quantitative PCR (ChIP-qPCR) | Diagenode',
'modified' => '2018-01-09 16:46:56',
'created' => '2014-12-11 00:22:08',
'ProductsApplication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '2',
'position' => '10',
'parent_id' => null,
'name' => 'エピジェネティクス・クロマチン解析',
'description' => '<div class="row">
<div class="large-12 columns">エピジェネティクス研究は、異なる転写パターン、遺伝子発現およびサイレンシングを引き起こすクロマチンの変化に対処します。<br /><br />クロマチンの主成分はDNA<span>およびヒストン蛋白質です。<span> </span></span>各ヒストンコア蛋白質(H2A<span>、</span>H2B<span>、</span>H3<span>および</span>H4<span>)の</span>2<span>つのコピーを</span>8<span>量体に組み込み、</span>DNA<span>で包んでヌクレオソームコアを形成させます。<span> </span></span>ヌクレオソームは、転写機械のDNA<span>への接近可能性および</span>クロマチン再構成因子を制御します。</div>
<div class="large-12 columns">
<p></p>
<p>クロマチン免疫沈降(ChIP<span>)は、関心対象の特定の蛋白質に対するゲノム結合部位の位置を解明するために使用される方法であり、遺伝子発現の制御に関する非常に貴重な洞察を提供します。<span> </span></span>ChIPは特定の抗原を含むクロマチン断片の選択的富化に関与します。 特定の蛋白質または蛋白質修飾を認識する抗体を使用して、特定の遺伝子座における抗原の相対存在量を決定します。</p>
<p>ChIP-seq<span>および</span>ChIP-qPCR<span>は、蛋白質</span>-DNA<span>結合部位の同定を可能にする技術です。</span></p>
<p> </p>
</div>
</div>',
'in_footer' => true,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'epigenetics-chromatin-study',
'meta_keywords' => 'Chromatin Immunoprecipitation Sequencing,ChIP-Seq,ChIP-seq grade antibodies,DNA purification,qPCR,Shearing of chromatin',
'meta_description' => 'Diagenodeは、ChIP-Seqおよびクロマチン免疫沈降(ChIP)専用キットと抗体を定量PCRアプリケーションと組み合わせて幅広く提供しています。',
'meta_title' => 'エピジェネティクス - クロマチン免疫沈降(ChIP)| Diagenode',
'modified' => '2018-01-11 09:58:47',
'created' => '2015-02-20 16:10:25',
'ProductsApplication' => array(
[maximum depth reached]
)
)
),
'Category' => array(
(int) 0 => array(
'id' => '61',
'position' => '2',
'parent_id' => '14',
'name' => 'Column-based DNA purification',
'description' => '',
'no_promo' => false,
'in_menu' => false,
'online' => true,
'tabular' => true,
'hide' => false,
'all_format' => false,
'is_antibody' => false,
'slug' => 'column-based-dna-purification',
'cookies_tag_id' => null,
'meta_keywords' => 'MicroChIP DiaPure columns,diagenode',
'meta_description' => 'Diagenode provides Column-based DNA purification for downstream analysis.',
'meta_title' => 'Column-based DNA purification | Diagenode',
'modified' => '2016-01-21 13:49:21',
'created' => '2015-07-08 10:31:04',
'ProductsCategory' => array(
[maximum depth reached]
),
'CookiesTag' => array([maximum depth reached])
)
),
'Document' => array(
(int) 0 => array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
[maximum depth reached]
)
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
'id' => '5033',
'name' => 'Trithorax regulates long-term memory in Drosophila through epigenetic maintenance of mushroom body metabolic state and translation capacity',
'authors' => 'Nicholas Raun et al.',
'description' => '<p><span>The role of epigenetics and chromatin in the maintenance of postmitotic neuronal cell identities is not well understood. Here, we show that the histone methyltransferase Trithorax (Trx) is required in postmitotic memory neurons of the </span><em>Drosophila</em><span><span> </span>mushroom body (MB) to enable their capacity for long-term memory (LTM), but not short-term memory (STM). Using MB-specific RNA-, ChIP-, and ATAC-sequencing, we find that Trx maintains homeostatic expression of several non-canonical MB-enriched transcripts, including the orphan nuclear receptor<span> </span></span><em>Hr51</em><span>, and the metabolic enzyme<span> </span></span><em>lactate dehydrogenase</em><span><span> </span>(</span><em>Ldh)</em><span>. Through these key targets, Trx facilitates a metabolic state characterized by high lactate levels in MBγ neurons. This metabolic state supports a high capacity for protein translation, a process that is essential for LTM, but not STM. These data suggest that Trx, a classic regulator of cell type specification during development, has additional functions in maintaining underappreciated aspects of postmitotic neuron identity, such as metabolic state. Our work supports a body of evidence suggesting that a high capacity for energy metabolism is an essential cell identity characteristic for neurons that mediate LTM.</span></p>',
'date' => '2025-01-27',
'pmid' => 'https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.3003004',
'doi' => 'https://doi.org/10.1371/journal.pbio.3003004',
'modified' => '2025-01-30 10:12:42',
'created' => '2025-01-30 10:12:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '4983',
'name' => 'Integrated multi-omics analysis of PBX1 in mouse adult neural stem- and progenitor cells identifies a transcriptional module that functionally links PBX1 to TCF3/4',
'authors' => 'Vera Laub et al.',
'description' => '<p><span>Developmental transcription factors act in networks, but how these networks achieve cell- and tissue specificity is still poorly understood. Here, we explored pre-B cell leukemia homeobox 1 (PBX1) in adult neurogenesis combining genomic, transcriptomic, and proteomic approaches. ChIP-seq analysis uncovered PBX1 binding to numerous genomic sites. Integration of PBX1 ChIP-seq with ATAC-seq data predicted interaction partners, which were subsequently validated by mass spectrometry. Whole transcriptome spatial RNA analysis revealed shared expression dynamics of </span><em>Pbx1</em><span><span> </span>and interacting factors. Among these were class I bHLH proteins TCF3 and TCF4. RNA-seq following<span> </span></span><em>Pbx1</em><span>,<span> </span></span><em>Tcf3</em><span><span> </span>or<span> </span></span><em>Tcf4</em><span><span> </span>knockdown identified proliferation- and differentiation associated genes as shared targets, while sphere formation assays following knockdown argued for functional cooperativity of PBX1 and TCF3 in progenitor cell proliferation. Notably, while physiological PBX1-TCF interaction has not yet been described, chromosomal translocation resulting in genomic<span> </span></span><em>TCF3::PBX1</em><span><span> </span>fusion characterizes a subtype of acute lymphoblastic leukemia. Introducing<span> </span></span><em>Pbx1</em><span><span> </span>into Nalm6 cells, a pre-B cell line expressing<span> </span></span><em>TCF3</em><span><span> </span>but lacking<span> </span></span><em>PBX1</em><span>, upregulated the leukemogenic genes<span> </span></span><em>BLK</em><span><span> </span>and<span> </span></span><em>NOTCH3</em><span>, arguing that functional PBX1-TCF cooperativity likely extends to hematopoiesis. Our study hence uncovers a transcriptional module orchestrating the balance between progenitor cell proliferation and differentiation in adult neurogenesis with potential implications for leukemia etiology.</span></p>',
'date' => '2024-10-08',
'pmid' => 'https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkae864/7815639',
'doi' => 'https://doi.org/10.1093/nar/gkae864',
'modified' => '2024-10-11 10:02:42',
'created' => '2024-10-11 10:02:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '4964',
'name' => 'A PRE loop at the dac locus acts as a topological chromatin structure that restricts and specifies enhancer–promoter communication',
'authors' => 'Denaud S. et al.',
'description' => '<p><span>Three-dimensional (3D) genome folding has a fundamental role in the regulation of developmental genes by facilitating or constraining chromatin interactions between </span><i>cis</i><span>-regulatory elements (CREs). Polycomb response elements (PREs) are a specific kind of CRE involved in the memory of transcriptional states in<span> </span></span><i>Drosophila melanogaster</i><span>. PREs act as nucleation sites for Polycomb group (PcG) proteins, which deposit the repressive histone mark H3K27me3, leading to the formation of a class of topologically associating domain (TAD) called a Polycomb domain. PREs can establish looping contacts that stabilize the gene repression of key developmental genes during development. However, the mechanism by which PRE loops fine-tune gene expression is unknown. Using clustered regularly interspaced short palindromic repeats and Cas9 genome engineering, we specifically perturbed PRE contacts or enhancer function and used complementary approaches including 4C-seq, Hi-C and Hi-M to analyze how chromatin architecture perturbation affects gene expression. Our results suggest that the PRE loop at the<span> </span></span><i>dac</i><span><span> </span>gene locus acts as a constitutive 3D chromatin scaffold during<span> </span></span><i>Drosophila</i><span><span> </span>development that forms independently of gene expression states and has a versatile function; it restricts enhancer–promoter communication and contributes to enhancer specificity.</span></p>',
'date' => '2024-08-16',
'pmid' => 'https://www.nature.com/articles/s41594-024-01375-7',
'doi' => 'https://doi.org/10.1038/s41594-024-01375-7',
'modified' => '2024-09-02 10:20:28',
'created' => '2024-09-02 10:20:28',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '4954',
'name' => 'A multiomic atlas of the aging hippocampus reveals molecular changes in response to environmental enrichment',
'authors' => 'Perez R. F. at al. ',
'description' => '<p><span>Aging involves the deterioration of organismal function, leading to the emergence of multiple pathologies. Environmental stimuli, including lifestyle, can influence the trajectory of this process and may be used as tools in the pursuit of healthy aging. To evaluate the role of epigenetic mechanisms in this context, we have generated bulk tissue and single cell multi-omic maps of the male mouse dorsal hippocampus in young and old animals exposed to environmental stimulation in the form of enriched environments. We present a molecular atlas of the aging process, highlighting two distinct axes, related to inflammation and to the dysregulation of mRNA metabolism, at the functional RNA and protein level. Additionally, we report the alteration of heterochromatin domains, including the loss of bivalent chromatin and the uncovering of a heterochromatin-switch phenomenon whereby constitutive heterochromatin loss is partially mitigated through gains in facultative heterochromatin. Notably, we observed the multi-omic reversal of a great number of aging-associated alterations in the context of environmental enrichment, which was particularly linked to glial and oligodendrocyte pathways. In conclusion, our work describes the epigenomic landscape of environmental stimulation in the context of aging and reveals how lifestyle intervention can lead to the multi-layered reversal of aging-associated decline.</span></p>',
'date' => '2024-07-16',
'pmid' => 'https://www.nature.com/articles/s41467-024-49608-z',
'doi' => 'https://doi.org/10.1038/s41467-024-49608-z',
'modified' => '2024-07-29 11:33:49',
'created' => '2024-07-29 11:33:49',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '4927',
'name' => 'Inflammatory stress-mediated chromatin changes underlie dysfunction in endothelial cells',
'authors' => 'Liu H. et al.',
'description' => '<p><span>Inflammatory stresses underlie endothelial dysfunction and contribute to the development of chronic cardiovascular disorders such as atherosclerosis and vascular fibrosis. The initial transcriptional response of endothelial cells to pro-inflammatory cytokines such as TNF-alpha is well established. However, very few studies uncover the effects of inflammatory stresses on chromatin architecture. We used integrative analysis of ATAC-seq and RNA-seq data to investigate chromatin alterations in human endothelial cells in response to TNF-alpha and febrile-range heat stress exposure. Multi-omics data analysis suggests a correlation between the transcription of stress-related genes and endothelial dysfunction drivers with chromatin regions exhibiting differential accessibility. Moreover, microscopy identified the dynamics in the nuclear organization, specifically, the changes in a subset of heterochromatic nucleoli-associated chromatin domains, the centromeres. Upon inflammatory stress exposure, the centromeres decreased association with nucleoli in a p38-dependent manner and increased the number of transcripts from pericentromeric regions. Overall, we provide two lines of evidence that suggest chromatin alterations in vascular endothelial cells during inflammatory stresses.</span></p>',
'date' => '2023-10-16',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10614786/',
'doi' => '10.1101/2023.10.11.561959',
'modified' => '2024-03-27 15:15:21',
'created' => '2024-03-27 15:15:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '4862',
'name' => 'Mutant FUS induces chromatin reorganization in the hippocampus andalters memory processes.',
'authors' => 'Tzeplaeff L. et al.',
'description' => '<p>Cytoplasmic mislocalization of the nuclear Fused in Sarcoma (FUS) protein is associated to amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Cytoplasmic FUS accumulation is recapitulated in the frontal cortex and spinal cord of heterozygous Fus mice. Yet, the mechanisms linking FUS mislocalization to hippocampal function and memory formation are still not characterized. Herein, we show that in these mice, the hippocampus paradoxically displays nuclear FUS accumulation. Multi-omic analyses showed that FUS binds to a set of genes characterized by the presence of an ETS/ELK-binding motifs, and involved in RNA metabolism, transcription, ribosome/mitochondria and chromatin organization. Importantly, hippocampal nuclei showed a decompaction of the neuronal chromatin at highly expressed genes and an inappropriate transcriptomic response was observed after spatial training of Fus mice. Furthermore, these mice lacked precision in a hippocampal-dependent spatial memory task and displayed decreased dendritic spine density. These studies shows that mutated FUS affects epigenetic regulation of the chromatin landscape in hippocampal neurons, which could participate in FTD/ALS pathogenic events. These data call for further investigation in the neurological phenotype of FUS-related diseases and open therapeutic strategies towards epigenetic drugs.</p>',
'date' => '2023-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/37327984',
'doi' => '10.1016/j.pneurobio.2023.102483',
'modified' => '2023-08-01 14:55:49',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '4733',
'name' => 'FXR inhibition may protect from SARS-CoV-2 infection by reducingACE2.',
'authors' => 'Brevini Teresa et al.',
'description' => '<p>Prevention of SARS-CoV-2 infection through the modulation of viral host receptors, such as ACE2, could represent a new chemoprophylactic approach for COVID-19 complementing vaccination. However, the mechanisms controlling ACE2 expression remain elusive. Here, we identify the farnesoid X receptor (FXR) as a direct regulator of ACE2 transcription in multiple COVID19-affected tissues, including the gastrointestinal and respiratory systems. We then use the over-the-counter compound z-guggulsterone (ZGG) and the off-patent drug ursodeoxycholic acid (UDCA) to reduce FXR signalling and downregulate ACE2 in human lung, cholangiocyte and intestinal organoids and in the corresponding tissues in mice and hamsters. We demonstrate that UDCA-mediated ACE2 downregulation reduces susceptibility to SARS-CoV-2 infection in vitro, in vivo and in human lungs and livers perfused ex situ. Furthermore, we illustrate that UDCA reduces ACE2 expression in the nasal epithelium in humans. Finally, we identify a correlation between UDCA treatment and positive clinical outcomes following SARS-CoV-2 infection using retrospective registry data, and confirm these findings in an independent validation cohort of liver transplant recipients. In conclusion, we identify a novel function of FXR in controlling ACE2 expression and provide evidence that modulation of this pathway could be beneficial for reducing SARS-CoV-2 infection, paving the road for future clinical trials.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36470304',
'doi' => '10.1038/s41586-022-05594-0',
'modified' => '2023-03-13 08:52:11',
'created' => '2023-02-28 12:19:11',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '4575',
'name' => 'Intranasal administration of Acinetobacter lwoffii in a murine model ofasthma induces IL-6-mediated protection associated with cecal microbiotachanges.',
'authors' => 'Alashkar A. B. et al.',
'description' => '<p>BACKGROUND: Early-life exposure to certain environmental bacteria including Acinetobacter lwoffii (AL) has been implicated in protection from chronic inflammatory diseases including asthma later in life. However, the underlying mechanisms at the immune-microbe interface remain largely unknown. METHODS: The effects of repeated intranasal AL exposure on local and systemic innate immune responses were investigated in wild-type and Il6 , Il10 , and Il17 mice exposed to ovalbumin-induced allergic airway inflammation. Those investigations were expanded by microbiome analyses. To assess for AL-associated changes in gene expression, the picture arising from animal data was supplemented by in vitro experiments of macrophage and T-cell responses, yielding expression and epigenetic data. RESULTS: The asthma preventive effect of AL was confirmed in the lung. Repeated intranasal AL administration triggered a proinflammatory immune response particularly characterized by elevated levels of IL-6, and consequently, IL-6 induced IL-10 production in CD4 T-cells. Both IL-6 and IL-10, but not IL-17, were required for asthma protection. AL had a profound impact on the gene regulatory landscape of CD4 T-cells which could be largely recapitulated by recombinant IL-6. AL administration also induced marked changes in the gastrointestinal microbiome but not in the lung microbiome. By comparing the effects on the microbiota according to mouse genotype and AL-treatment status, we have identified microbial taxa that were associated with either disease protection or activity. CONCLUSION: These experiments provide a novel mechanism of Acinetobacter lwoffii-induced asthma protection operating through IL-6-mediated epigenetic activation of IL-10 production and with associated effects on the intestinal microbiome.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36458896',
'doi' => '10.1111/all.15606',
'modified' => '2023-04-11 10:23:07',
'created' => '2023-02-21 09:59:46',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 8 => array(
'id' => '4836',
'name' => 'Caffeine intake exerts dual genome-wide effects on hippocampal metabolismand learning-dependent transcription.',
'authors' => 'Paiva I. et al.',
'description' => '<p>Caffeine is the most widely consumed psychoactive substance in the world. Strikingly, the molecular pathways engaged by its regular consumption remain unclear. We herein addressed the mechanisms associated with habitual (chronic) caffeine consumption in the mouse hippocampus using untargeted orthogonal omics techniques. Our results revealed that chronic caffeine exerts concerted pleiotropic effects in the hippocampus at the epigenomic, proteomic, and metabolomic levels. Caffeine lowered metabolism-related processes (e.g., at the level of metabolomics and gene expression) in bulk tissue, while it induced neuron-specific epigenetic changes at synaptic transmission/plasticity-related genes and increased experience-driven transcriptional activity. Altogether, these findings suggest that regular caffeine intake improves the signal-to-noise ratio during information encoding, in part through fine-tuning of metabolic genes, while boosting the salience of information processing during learning in neuronal circuits.</p>',
'date' => '2022-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35536645',
'doi' => '10.1172/JCI149371',
'modified' => '2023-08-01 13:52:29',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 9 => array(
'id' => '4220',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in Mouse Prostate Cancer Xenografts',
'authors' => 'Sanchez A. et al.',
'description' => '<p><strong class="sub-title">Background/aim:<span> </span></strong>Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo.</p>
<p><strong class="sub-title">Materials and methods:<span> </span></strong>Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR.</p>
<p><strong class="sub-title">Results:<span> </span></strong>JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression.</p>
<p><strong class="sub-title">Conclusion:<span> </span></strong>JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-05-01',
'pmid' => 'https://pubmed.ncbi.nlm.nih.gov/35430567/',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-04-21 11:54:21',
'created' => '2022-04-21 11:54:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 10 => array(
'id' => '4409',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in MouseProstate Cancer Xenografts.',
'authors' => 'Sanchez A. et al.',
'description' => '<p>BACKGROUND/AIM: Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo. MATERIALS AND METHODS: Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR. RESULTS: JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression. CONCLUSION: JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35430567',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-08-11 15:11:58',
'created' => '2022-08-11 12:14:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 11 => array(
'id' => '4233',
'name' => 'Autocrine Vitamin D-signaling switches off pro-inflammatory programsof Th1 cells',
'authors' => 'Chauss D.et al.',
'description' => '<p>The molecular mechanisms governing orderly shutdown and retraction of CD4+ T helper (Th)1 responses remain poorly understood. Here, we show that complement triggers contraction of Th1 responses by inducing intrinsic expression of the vitamin D (VitD) receptor (VDR) and the VitD-activating enzyme CYP27B1, permitting T cells to both activate and respond to VitD. VitD then initiated transition from pro-inflammatory IFN-γ + Th1 cells to suppressive IL-10+ cells. This process was primed by dynamic changes in the epigenetic landscape of CD4+ T cells, generating super-enhancers and recruiting several transcription factors, notably c-JUN, STAT3 and BACH2, which together with VDR shaped the transcriptional response to VitD. Accordingly, VitD did not induce IL-10 in cells with dysfunctional BACH2 or STAT3. Bronchoalveolar lavage fluid CD4+ T cells of COVID-19 patients were Th1-skewed and showed de-repression of genes down-regulated by VitD, either from lack of substrate (VitD deficiency) and/or abnormal regulation of this system.</p>',
'date' => '2021-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/34764490',
'doi' => '10.1038/s41590-021-01080-3',
'modified' => '2022-05-19 16:57:27',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 12 => array(
'id' => '4351',
'name' => 'Essential role of a ThPOK autoregulatory loop in the maintenance ofmature CD4 T cell identity and function.',
'authors' => 'Basu Jayati et al.',
'description' => '<p>The transcription factor ThPOK (encoded by the Zbtb7b gene) controls homeostasis and differentiation of mature helper T cells, while opposing their differentiation to CD4 intraepithelial lymphocytes (IELs) in the intestinal mucosa. Thus CD4 IEL differentiation requires ThPOK transcriptional repression via reactivation of the ThPOK transcriptional silencer element (Sil). In the present study, we describe a new autoregulatory loop whereby ThPOK binds to the Sil to maintain its own long-term expression in CD4 T cells. Disruption of this loop in vivo prevents persistent ThPOK expression, leads to genome-wide changes in chromatin accessibility and derepresses the colonic regulatory T (T) cell gene expression signature. This promotes selective differentiation of naive CD4 T cells into GITRPD-1CD25 (Triple) T cells and conversion to CD4 IELs in the gut, thereby providing dominant protection from colitis. Hence, the ThPOK autoregulatory loop represents a key mechanism to physiologically control ThPOK expression and T cell differentiation in the gut, with potential therapeutic relevance.</p>',
'date' => '2021-08-01',
'pmid' => 'https://doi.org/10.1038%2Fs41590-021-00980-8',
'doi' => '10.1038/s41590-021-00980-8',
'modified' => '2022-06-22 12:32:59',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 13 => array(
'id' => '4127',
'name' => 'The histone modification H3K4me3 is altered at the locus in Alzheimer'sdisease brain.',
'authors' => 'Smith, Adam et al.',
'description' => '<p>Several epigenome-wide association studies of DNA methylation have highlighted altered DNA methylation in the gene in Alzheimer's disease (AD) brain samples. However, no study has specifically examined histone modifications in the disease. We use chromatin immunoprecipitation-qPCR to quantify tri-methylation at histone 3 lysine 4 (H3K4me3) and 27 (H3K27me3) in the gene in entorhinal cortex from donors with high (n = 59) or low (n = 29) Alzheimer's disease pathology. We demonstrate decreased levels of H3K4me3, a marker of active gene transcription, with no change in H3K27me3, a marker of inactive genes. H3K4me3 is negatively correlated with DNA methylation in specific regions of the gene. Our study suggests that the gene shows altered epigenetic marks indicative of reduced gene activation in Alzheimer's disease.</p>',
'date' => '2021-02-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/33815817',
'doi' => '10.2144/fsoa-2020-0161',
'modified' => '2021-12-07 10:16:08',
'created' => '2021-12-06 15:53:19',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 14 => array(
'id' => '4062',
'name' => 'Digging Deeper into Breast Cancer Epigenetics: Insights from ChemicalInhibition of Histone Acetyltransferase TIP60 .',
'authors' => 'Idrissou, Mouhamed and Lebert, Andre and Boisnier, Tiphanie and Sanchez,Anna and Houfaf Khoufaf, Fatma Zohra and Penault-Llorca, Frédérique andBignon, Yves-Jean and Bernard-Gallon, Dominique',
'description' => '<p>Breast cancer is often sporadic due to several factors. Among them, the deregulation of epigenetic proteins may be involved. TIP60 or KAT5 is an acetyltransferase that regulates gene transcription through the chromatin structure. This pleiotropic protein acts in several cellular pathways by acetylating proteins. RNA and protein expressions of TIP60 were shown to decrease in some breast cancer subtypes, particularly in triple-negative breast cancer (TNBC), where a low expression of TIP60 was exhibited compared with luminal subtypes. In this study, the inhibition of the residual activity of TIP60 in breast cancer cell lines was investigated by using two chemical inhibitors, TH1834 and NU9056, first on the acetylation of the specific target, lysine 4 of histone 3 (H3K4) by immunoblotting, and second, by chromatin immunoprecipitation (ChIP)-qPCR (-quantitative Polymerase Chain Reaction). Subsequently, significant decreases or a trend toward decrease of H3K4ac in the different chromatin compartments were observed. In addition, the expression of 48 human nuclear receptors was studied with TaqMan Low-Density Array in these breast cancer cell lines treated with TIP60 inhibitors. The statistical analysis allowed us to comprehensively characterize the androgen receptor and receptors in TNBC cell lines after TH1834 or NU9056 treatment. The understanding of the residual activity of TIP60 in the evolution of breast cancer might be a major asset in the fight against this disease, and could allow TIP60 to be used as a biomarker or therapeutic target for breast cancer progression in the future.</p>',
'date' => '2020-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/32960142',
'doi' => '10.1089/omi.2020.0104',
'modified' => '2021-02-19 17:39:52',
'created' => '2021-02-18 10:21:53',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 15 => array(
'id' => '3969',
'name' => 'Delineating the early transcriptional specification of the mammalian trachea and esophagus.',
'authors' => 'Kuwahara A, Lewis AE, Coombes C, Leung FS, Percharde M, Bush JO',
'description' => '<p>The genome-scale transcriptional programs that specify the mammalian trachea and esophagus are unknown. Though NKX2-1 and SOX2 are hypothesized to be co-repressive master regulators of tracheoesophageal fates, this is untested at a whole transcriptomic scale and their downstream networks remain unidentified. By combining single-cell RNA-sequencing with bulk RNA-sequencing of mutants and NKX2-1 ChIP-sequencing in mouse embryos, we delineate the NKX2-1 transcriptional program in tracheoesophageal specification, and discover that the majority of the tracheal and esophageal transcriptome is NKX2-1 independent. To decouple the NKX2-1 transcriptional program from regulation by SOX2, we interrogate the expression of newly-identified tracheal and esophageal markers in / compound mutants. Finally, we discover that NKX2-1 binds directly to and and regulates their expression to control mesenchymal specification to cartilage and smooth muscle, coupling epithelial identity with mesenchymal specification. These findings create a new framework for understanding early tracheoesophageal fate specification at the genome-wide level.</p>',
'date' => '2020-06-09',
'pmid' => 'http://www.pubmed.gov/32515350',
'doi' => '10.7554/eLife.55526',
'modified' => '2020-08-12 09:32:02',
'created' => '2020-08-10 12:12:25',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 16 => array(
'id' => '3845',
'name' => 'Combinatorial action of NF-Y and TALE at embryonic enhancers defines distinct gene expression programs during zygotic genome activation in zebrafish.',
'authors' => 'Stanney W, Ladam F, Donaldson IJ, Parsons TJ, Maehr R, Bobola N, Sagerström CG',
'description' => '<p>Animal embryogenesis is initiated by maternal factors, but zygotic genome activation (ZGA) shifts regulatory control to the embryo during blastula stages. ZGA is thought to be mediated by maternally provided transcription factors (TFs), but few such TFs have been identified in vertebrates. Here we report that NF-Y and TALE TFs bind zebrafish genomic elements associated with developmental control genes already at ZGA. In particular, co-regulation by NF-Y and TALE is associated with broadly acting genes involved in transcriptional control, while regulation by either NF-Y or TALE defines genes in specific developmental processes, such that NF-Y controls a cilia gene expression program while TALE controls expression of hox genes. We also demonstrate that NF-Y and TALE-occupied genomic elements function as enhancers during embryogenesis. We conclude that combinatorial use of NF-Y and TALE at developmental enhancers permits the establishment of distinct gene expression programs at zebrafish ZGA.</p>',
'date' => '2019-12-17',
'pmid' => 'http://www.pubmed.gov/31862379',
'doi' => '10.1016/j.ydbio.2019.12.003',
'modified' => '2020-02-20 11:13:27',
'created' => '2020-02-13 10:02:44',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 17 => array(
'id' => '3744',
'name' => 'Whsc1 links pluripotency exit with mesendoderm specification.',
'authors' => 'Tian TV, Di Stefano B, Stik G, Vila-Casadesús M, Sardina JL, Vidal E, Dasti A, Segura-Morales C, De Andrés-Aguayo L, Gómez A, Goldmann J, Jaenisch R, Graf T',
'description' => '<p>How pluripotent stem cells differentiate into the main germ layers is a key question of developmental biology. Here, we show that the chromatin-related factor Whsc1 (also known as Nsd2 and MMSET) has a dual role in pluripotency exit and germ layer specification of embryonic stem cells. On induction of differentiation, a proportion of Whsc1-depleted embryonic stem cells remain entrapped in a pluripotent state and fail to form mesendoderm, although they are still capable of generating neuroectoderm. These functions of Whsc1 are independent of its methyltransferase activity. Whsc1 binds to enhancers of the mesendodermal regulators Gata4, T (Brachyury), Gata6 and Foxa2, together with Brd4, and activates the expression of these genes. Depleting each of these regulators also delays pluripotency exit, suggesting that they mediate the effects observed with Whsc1. Our data indicate that Whsc1 links silencing of the pluripotency regulatory network with activation of mesendoderm lineages.</p>',
'date' => '2019-07-01',
'pmid' => 'http://www.pubmed.gov/31235934',
'doi' => '10.1038/s41556-019-0342-1',
'modified' => '2019-08-06 16:35:35',
'created' => '2019-07-31 13:35:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 18 => array(
'id' => '3639',
'name' => 'Spatial confinement downsizes the inflammatory response of macrophages.',
'authors' => 'Jain N, Vogel V',
'description' => '<p>Macrophages respond to chemical/metabolic and physical stimuli, but their effects cannot be readily decoupled in vivo during pro-inflammatory activation. Here, we show that preventing macrophage spreading by spatial confinement, as imposed by micropatterning, microporous substrates or cell crowding, suppresses late lipopolysaccharide (LPS)-activated transcriptional programs (biomarkers IL-6, CXCL9, IL-1β, and iNOS) by mechanomodulating chromatin compaction and epigenetic alterations (HDAC3 levels and H3K36-dimethylation). Mechanistically, confinement reduces actin polymerization, thereby lowers the LPS-stimulated nuclear translocation of MRTF-A. This lowers the activity of the MRTF-A-SRF complex and subsequently downregulates the inflammatory response, as confirmed by chromatin immunoprecipitation coupled with quantitative PCR and RNA sequencing analysis. Confinement thus downregulates pro-inflammatory cytokine secretion and, well before any activation processes, the phagocytic potential of macrophages. Contrarily, early events, including activation of the LPS receptor TLR4, and downstream NF-κB and IRF3 signalling and hence the expression of early LPS-responsive genes were marginally affected by confinement. These findings have broad implications in the context of mechanobiology, inflammation and immunology, as well as in tissue engineering and regenerative medicine.</p>',
'date' => '2018-12-01',
'pmid' => 'http://www.pubmed.gov/30349032',
'doi' => '10.1038/s41563-018-0190-6',
'modified' => '2019-06-07 10:23:26',
'created' => '2019-06-06 12:11:18',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 19 => array(
'id' => '3379',
'name' => 'SIRT1-dependent epigenetic regulation of H3 and H4 histone acetylation in human breast cancer',
'authors' => 'Khaldoun Rifaï et al.',
'description' => '<p>Breast cancer is the most frequently diagnosed malignancy in women worldwide. It is well established that the complexity of carcinogenesis involves profound epigenetic deregulations that contribute to the tumorigenesis process. Deregulated H3 and H4 acetylated histone marks are amongst those alterations. Sirtuin-1 (SIRT1) is a class-III histone deacetylase deeply involved in apoptosis, genomic stability, gene expression regulation and breast tumorigenesis. However, the underlying molecular mechanism by which SIRT1 regulates H3 and H4 acetylated marks, and consequently cancer-related gene expression in breast cancer, remains uncharacterized. In this study, we elucidated SIRT1 epigenetic role and analyzed the link between the latter and histones H3 and H4 epigenetic marks in all 5 molecular subtypes of breast cancer. Using a cohort of 135 human breast tumors and their matched normal tissues, as well as 5 human-derived cell lines, we identified H3k4ac as a new prime target of SIRT1 in breast cancer. We also uncovered an inverse correlation between SIRT1 and the 3 epigenetic marks H3k4ac, H3k9ac and H4k16ac expression patterns. We showed that SIRT1 modulates the acetylation patterns of histones H3 and H4 in breast cancer. Moreover, SIRT1 regulates its H3 acetylated targets in a subtype-specific manner. Furthermore, SIRT1 siRNA-mediated knockdown increases histone acetylation levels at 6 breast cancer-related gene promoters: <em>AR</em>, <em>BRCA1</em>, <em>ERS1</em>, <em>ERS2</em>, <em>EZH2</em> and <em>EP300</em>. In summary, this report characterizes for the first time the epigenetic behavior of SIRT1 in human breast carcinoma. These novel findings point to a potential use of SIRT1 as an epigenetic therapeutic target in breast cancer.</p>',
'date' => '2018-07-17',
'pmid' => 'http://www.oncotarget.com/index.php?journal=oncotarget&page=article&op=view&path[]=25771&path[]=80619',
'doi' => '',
'modified' => '2018-08-09 10:47:58',
'created' => '2018-07-26 12:02:12',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 20 => array(
'id' => '3595',
'name' => 'Reciprocal signalling by Notch-Collagen V-CALCR retains muscle stem cells in their niche.',
'authors' => 'Baghdadi MB, Castel D, Machado L, Fukada SI, Birk DE, Relaix F, Tajbakhsh S, Mourikis P',
'description' => '<p>The cell microenvironment, which is critical for stem cell maintenance, contains both cellular and non-cellular components, including secreted growth factors and the extracellular matrix. Although Notch and other signalling pathways have previously been reported to regulate quiescence of stem cells, the composition and source of molecules that maintain the stem cell niche remain largely unknown. Here we show that adult muscle satellite (stem) cells in mice produce extracellular matrix collagens to maintain quiescence in a cell-autonomous manner. Using chromatin immunoprecipitation followed by sequencing, we identified NOTCH1/RBPJ-bound regulatory elements adjacent to specific collagen genes, the expression of which is deregulated in Notch-mutant mice. Moreover, we show that Collagen V (COLV) produced by satellite cells is a critical component of the quiescent niche, as depletion of COLV by conditional deletion of the Col5a1 gene leads to anomalous cell cycle entry and gradual diminution of the stem cell pool. Notably, the interaction of COLV with satellite cells is mediated by the Calcitonin receptor, for which COLV acts as a surrogate local ligand. Systemic administration of a calcitonin derivative is sufficient to rescue the quiescence and self-renewal defects found in COLV-null satellite cells. This study reveals a Notch-COLV-Calcitonin receptor signalling cascade that maintains satellite cells in a quiescent state in a cell-autonomous fashion, and raises the possibility that similar reciprocal mechanisms act in diverse stem cell populations.</p>',
'date' => '2018-05-23',
'pmid' => 'http://www.pubmed.gov/29795344',
'doi' => '10.1038/s41586-018-0144-9',
'modified' => '2019-04-17 15:12:55',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 21 => array(
'id' => '3589',
'name' => 'A new metabolic gene signature in prostate cancer regulated by JMJD3 and EZH2.',
'authors' => 'Daures M, Idrissou M, Judes G, Rifaï K, Penault-Llorca F, Bignon YJ, Guy L, Bernard-Gallon D',
'description' => '<p>Histone methylation is essential for gene expression control. Trimethylated lysine 27 of histone 3 (H3K27me3) is controlled by the balance between the activities of JMJD3 demethylase and EZH2 methyltransferase. This epigenetic mark has been shown to be deregulated in prostate cancer, and evidence shows H3K27me3 enrichment on gene promoters in prostate cancer. To study the impact of this enrichment, a transcriptomic analysis with TaqMan Low Density Array (TLDA) of several genes was studied on prostate biopsies divided into three clinical grades: normal ( = 23) and two tumor groups that differed in their aggressiveness (Gleason score ≤ 7 ( = 20) and >7 ( = 19)). ANOVA demonstrated that expression of the gene set was upregulated in tumors and correlated with Gleason score, thus discriminating between the three clinical groups. Six genes involved in key cellular processes stood out: , , , , and . Chromatin immunoprecipitation demonstrated collocation of EZH2 and JMJD3 on gene promoters that was dependent on disease stage. Gene set expression was also evaluated on prostate cancer cell lines (DU 145, PC-3 and LNCaP) treated with an inhibitor of JMJD3 (GSK-J4) or EZH2 (DZNeP) to study their involvement in gene regulation. Results showed a difference in GSK-J4 sensitivity under PTEN status of cell lines and an opposite gene expression profile according to androgen status of cells. In summary, our data describe the impacts of JMJD3 and EZH2 on a new gene signature involved in prostate cancer that may help identify diagnostic and therapeutic targets in prostate cancer.</p>',
'date' => '2018-05-04',
'pmid' => 'http://www.pubmed.gov/29805743',
'doi' => '10.18632/oncotarget.25182',
'modified' => '2019-04-17 15:21:33',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 22 => array(
'id' => '3477',
'name' => 'Contrasting epigenetic states of heterochromatin in the different types of mouse pluripotent stem cells.',
'authors' => 'Tosolini M, Brochard V, Adenot P, Chebrout M, Grillo G, Navia V, Beaujean N, Francastel C, Bonnet-Garnier A, Jouneau A',
'description' => '<p>Mouse embryonic stem cells (ESCs) and epiblast stem cells (EpiSCs) represent naive and primed pluripotency states, respectively, and are maintained in vitro by specific signalling pathways. Furthermore, ESCs cultured in serum-free medium with two kinase inhibitors (2i-ESCs) are thought to be the ground naïve pluripotent state. Here, we present a comparative study of the epigenetic and transcriptional states of pericentromeric heterochromatin satellite sequences found in these pluripotent states. We show that 2i-ESCs are distinguished from other pluripotent cells by a prominent enrichment in H3K27me3 and low levels of DNA methylation at pericentromeric heterochromatin. In contrast, serum-containing ESCs exhibit higher levels of major satellite repeat transcription, which is lower in 2i-ESCs and even more repressed in primed EpiSCs. Removal of either DNA methylation or H3K9me3 at PCH in 2i-ESCs leads to enhanced deposition of H3K27me3 with few changes in satellite transcript levels. In contrast, their removal in EpiSCs does not lead to deposition of H3K27me3 but rather removes transcriptional repression. Altogether, our data show that the epigenetic state of PCH is modified during transition from naive to primed pluripotency states towards a more repressive state, which tightly represses the transcription of satellite repeats.</p>',
'date' => '2018-04-10',
'pmid' => 'http://www.pubmed.gov/29636490',
'doi' => '10.1038/s41598-018-23822-4',
'modified' => '2019-02-15 20:26:34',
'created' => '2019-02-14 15:01:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 23 => array(
'id' => '3324',
'name' => 'Isl1 mediates mesenchymal expansion in the developing external genitalia via regulation of Bmp4, Fgf10 and Wnt5a',
'authors' => 'Ching S.T. et al.',
'description' => '<p>Genital malformations are among the most common human birth defects, and both genetic and environmental factors can contribute to these malformations. Development of the external genitalia in mammals relies on complex signaling networks, and disruption of these signaling pathways can lead to genital defects. Islet-1 (ISL1), a member of the LIM/Homeobox family of transcription factors, has been identified as a major susceptibility gene for classic bladder exstrophy in humans, a common form of the bladder exstrophy-epispadias complex (BEEC), and is implicated in a role in urinary tract development. We report that deletion of Isl1 from the genital mesenchyme in mice led to hypoplasia of the genital tubercle and prepuce, with an ectopic urethral opening and epispadias-like phenotype. These mice also developed hydroureter and hydronephrosis. Identification of ISL1 transcriptional targets via ChIP-Seq and expression analyses revealed that Isl1 regulates several important signaling pathways during embryonic genital development, including the BMP, WNT, and FGF cascades. An essential function of Isl1 during development of the external genitalia is to induce Bmp4-mediated apoptosis in the genital mesenchyme. Together, these studies demonstrate that Isl1 plays a critical role during development of the external genitalia and forms the basis for a greater understanding of the molecular mechanisms underlying the pathogenesis of BEEC and urinary tract defects in humans.</p>',
'date' => '2018-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/29126155',
'doi' => '',
'modified' => '2018-02-06 09:25:22',
'created' => '2018-02-06 09:25:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 24 => array(
'id' => '3322',
'name' => 'In Situ Fixation Redefines Quiescence and Early Activation of Skeletal Muscle Stem Cells',
'authors' => 'Machado L. et al.',
'description' => '<div class="abstract">
<h2 class="sectionTitle" tabindex="0">Summary</h2>
<div class="content">
<p>State of the art techniques have been developed to isolate and analyze cells from various tissues, aiming to capture their <em>in vivo</em> state. However, the majority of cell isolation protocols involve lengthy mechanical and enzymatic dissociation steps followed by flow cytometry, exposing cells to stress and disrupting their physiological niche. Focusing on adult skeletal muscle stem cells, we have developed a protocol that circumvents the impact of isolation procedures and captures cells in their native quiescent state. We show that current isolation protocols induce major transcriptional changes accompanied by specific histone modifications while having negligible effects on DNA methylation. In addition to proposing a protocol to avoid isolation-induced artifacts, our study reveals previously undetected quiescence and early activation genes of potential biological interest.</p>
</div>
</div>',
'date' => '2017-11-14',
'pmid' => 'http://www.cell.com/cell-reports/abstract/S2211-1247(17)31543-7',
'doi' => '',
'modified' => '2022-05-19 16:11:43',
'created' => '2018-02-02 16:36:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 25 => array(
'id' => '3263',
'name' => 'The RPMI-1640 vitamin mixture promotes bovine blastocyst development in vitro and downregulates gene expression of TXNIP with epigenetic modification of associated histones',
'authors' => 'Ikeda S.et al.',
'description' => '<p>Diverse environmental conditions surrounding preimplantation embryos, including available nutrients, affect their metabolism and development in both short- and long-term manner. Thioredoxin-interacting protein (TXNIP) is a possible marker for preimplantation stress that is implicated in in vitro fertilization- (IVF) induced long-term DOHaD effects. B vitamins, as participants in one-carbon metabolism, may affect preimplantation embryos by epigenetic alterations of metabolically and developmentally important genes. In vitro-produced bovine embryos were cultured with or without Roswell Park Memorial Institute 1640 vitamin mixture, containing B vitamins and B vitamin-like substances, from day 3 after IVF and we evaluated blastocyst development and TXNIP messenger RNA (mRNA) expression in the blastocysts by reverse transcription-quantitative polymerase chain reaction. The degree of trimethylation of histone H3 lysine 27 (H3K27me3) at TXNIP promoter was examined semi-quantitatively by chromatin immunoprecipitation polymerase chain reaction. Total H3K27me3 were also compared between the groups by Western blot analysis. The vitamin treatment significantly increased the rates of blastocyst development (P<0.05) and their hatching (P<0.001) from the zona pellucida by day 8. The mRNA expression of TXNIP was lower (P<0.01) in blastocysts in the vitamin-mixture-treated group concomitant with higher (P<0.05) level of H3K27me3 of its promoter compared with the control group. The total H3K27me3 in the vitamin-mixture-treated group was also higher (P<0.01) than that in the control group. The epigenetic control of genes related to important metabolic processes during the periconceptional period by nutritional conditions in utero and/or in vitro may have possible implication for the developmental programming during this period that may impact the welfare and production traits of farm animals.</p>',
'date' => '2017-08-02',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28764817',
'doi' => '',
'modified' => '2017-10-09 15:59:51',
'created' => '2017-10-09 15:59:51',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 26 => array(
'id' => '3193',
'name' => 'Global analysis of H3K27me3 as an epigenetic marker in prostate cancer progression',
'authors' => 'Ngollo M. et al.',
'description' => '<div class="">
<h4>BACKGROUND:</h4>
<p><abstracttext label="BACKGROUND" nlmcategory="BACKGROUND">H3K27me3 histone marks shape the inhibition of gene transcription. In prostate cancer, the deregulation of H3K27me3 marks might play a role in prostate tumor progression.</abstracttext></p>
<h4>METHODS:</h4>
<p><abstracttext label="METHODS" nlmcategory="METHODS">We investigated genome-wide H3K27me3 histone methylation profile using chromatin immunoprecipitation (ChIP) and 2X400K promoter microarrays to identify differentially-enriched regions in biopsy samples from prostate cancer patients. H3K27me3 marks were assessed in 34 prostate tumors: 11 with Gleason score > 7 (GS > 7), 10 with Gleason score ≤ 7 (GS ≤ 7), and 13 morphologically normal prostate samples.</abstracttext></p>
<h4>RESULTS:</h4>
<p><abstracttext label="RESULTS" nlmcategory="RESULTS">Here, H3K27me3 profiling identified an average of 386 enriched-genes on promoter regions in healthy control group versus 545 genes in GS ≤ 7 and 748 genes in GS > 7 group. We then ran a factorial discriminant analysis (FDA) and compared the enriched genes in prostate-tumor biopsies and normal biopsies using ANOVA to identify significantly differentially-enriched genes. The analysis identified ALG5, EXOSC8, CBX1, GRID2, GRIN3B, ING3, MYO1D, NPHP3-AS1, MSH6, FBXO11, SND1, SPATS2, TENM4 and TRA2A genes. These genes are possibly associated with prostate cancer. Notably, the H3K27me3 histone mark emerged as a novel regulatory mechanism in poor-prognosis prostate cancer.</abstracttext></p>
<h4>CONCLUSIONS:</h4>
<p><abstracttext label="CONCLUSIONS" nlmcategory="CONCLUSIONS">Our findings point to epigenetic mark H3K27me3 as an important event in prostate carcinogenesis and progression. The results reported here provide new molecular insights into the pathogenesis of prostate cancer.</abstracttext></p>
</div>',
'date' => '2017-04-12',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28403887',
'doi' => '',
'modified' => '2017-06-19 14:07:35',
'created' => '2017-06-19 14:05:03',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 27 => array(
'id' => '3156',
'name' => 'Crebbp loss cooperates with Bcl2 over-expression to promote lymphoma in mice',
'authors' => 'Idoia García-Ramírez, Saber Tadros, Inés González-Herrero, Alberto Martín-Lorenzo, Guillermo Rodríguez-Hernández, Dalia Moore, Lucía Ruiz-Roca, Oscar Blanco, Diego Alonso-López, Javier De Las Rivas, Keenan Hartert, Romain Duval, David Klinkebiel, Martin B',
'description' => '<p><span>CREBBP is targeted by inactivating mutations in follicular lymphoma (FL) and diffuse large B-cell lymphoma (DLBCL). Here, we provide evidence from transgenic mouse models that Crebbp deletion results in deficits in B-cell development and can cooperate with Bcl2 over-expression to promote B-cell lymphoma. Through transcriptional and epigenetic profiling of these B-cells we found that Crebbp inactivation was associated with broad transcriptional alterations, but no changes in the patterns of histone acetylation at the proximal regulatory regions of these genes. However, B-cells with Crebbp inactivation showed high expression of Myc and patterns of altered histone acetylation that were localized to intragenic regions, enriched for Myc DNA binding motifs, and showed Myc binding. Through the analysis of CREBBP mutations from a large cohort of primary human FL and DLBCL, we show a significant difference in the spectrum of CREBBP mutations in these two diseases, with higher frequencies of nonsense/frameshift mutations in DLBCL compared to FL. Together our data therefore provide important links between Crebbp inactivation and Bcl2 dependence, and show a role for Crebbp inactivation in the induction of Myc expression. We suggest this may parallel the role of CREBBP frameshift/nonsense mutations in DLBCL that result in loss of the protein, but may contrast the role of missense mutations in the lysine acetyltransferase domain that are more frequently observed in FL and yield an inactive protein.</span></p>',
'date' => '2017-03-05',
'pmid' => 'http://www.bloodjournal.org/content/early/2017/03/13/blood-2016-08-733469?sso-checked=true',
'doi' => 'https://doi.org/10.1182/blood-2016-08-733469',
'modified' => '2017-05-11 11:17:42',
'created' => '2017-04-10 07:56:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 28 => array(
'id' => '3151',
'name' => 'Aorta macrophage inflammatory and epigenetic changes in a murine model of obstructive sleep apnea: Potential role of CD36.',
'authors' => 'Cortese R. et al.',
'description' => '<p>Obstructive sleep apnea (OSA) affects 8-10% of the population, is characterized by chronic intermittent hypoxia (CIH), and causally associates with cardiovascular morbidities. In CIH-exposed mice, closely mimicking the chronicity of human OSA, increased accumulation and proliferation of pro-inflammatory metabolic M1-like macrophages highly expressing CD36, emerged in aorta. Transcriptomic and MeDIP-seq approaches identified activation of pro-atherogenic pathways involving a complex interplay of histone modifications in functionally-relevant biological pathways, such as inflammation and oxidative stress in aorta macrophages. Discontinuation of CIH did not elicit significant improvements in aorta wall macrophage phenotype. However, CIH-induced aorta changes were absent in CD36 knockout mice, Our results provide mechanistic insights showing that CIH exposures during sleep in absence of concurrent pro-atherogenic settings (i.e., genetic propensity or dietary manipulation) lead to the recruitment of CD36(+)<sup>high</sup> macrophages to the aortic wall and trigger atherogenesis. Furthermore, long-term CIH-induced changes may not be reversible with usual OSA treatment.</p>',
'date' => '2017-02-27',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28240319',
'doi' => '',
'modified' => '2017-03-28 09:16:02',
'created' => '2017-03-28 09:16:02',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 29 => array(
'id' => '3029',
'name' => 'Loss of Ezh2 synergizes with JAK2-V617F in initiating myeloproliferative neoplasms and promoting myelofibrosis',
'authors' => 'Shimizu T et al.',
'description' => '<p>Myeloproliferative neoplasm (MPN) patients frequently show co-occurrence of <em>JAK2</em>-V617F and mutations in epigenetic regulator genes, including <em>EZH2</em>. In this study, we show that <em>JAK2</em>-V617F and loss of <em>Ezh2</em> in hematopoietic cells contribute synergistically to the development of MPN. The MPN phenotype induced by <em>JAK2</em>-V617F was accentuated in <em>JAK2</em>-V617F<em>;Ezh2<sup>−/−</sup></em> mice, resulting in very high platelet and neutrophil counts, more advanced myelofibrosis, and reduced survival. These mice also displayed expansion of the stem cell and progenitor cell compartments and a shift of differentiation toward megakaryopoiesis at the expense of erythropoiesis. Single cell limiting dilution transplantation with bone marrow from <em>JAK2</em>-V617F<em>;Ezh2<sup>+/−</sup></em> mice showed increased reconstitution and MPN disease initiation potential compared with <em>JAK2</em>-V617F alone. RNA sequencing in <em>Ezh2</em>-deficient hematopoietic stem cells (HSCs) and megakaryocytic erythroid progenitors identified highly up-regulated genes, including <em>Lin28b</em> and <em>Hmga2</em>, and chromatin immunoprecipitation (ChIP)–quantitative PCR (qPCR) analysis of their promoters revealed decreased H3K27me3 deposition. Forced expression of <em>Hmga2</em> resulted in increased chimerism and platelet counts in recipients of retrovirally transduced HSCs. <em>JAK2</em>-V617F–expressing mice treated with an Ezh2 inhibitor showed higher platelet counts than vehicle controls. Our data support the proposed tumor suppressor function of <em>EZH2</em> in patients with MPN and call for caution when considering using Ezh2 inhibitors in MPN.</p>',
'date' => '2016-07-11',
'pmid' => 'http://jem.rupress.org/content/213/8/1479.short?rss=1',
'doi' => '',
'modified' => '2016-09-08 15:24:48',
'created' => '2016-09-08 15:24:48',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 30 => array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
[maximum depth reached]
)
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
'id' => '174',
'name' => 'MicroChIP DiaPure columns SDS GB en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-GB-en-1_0.pdf',
'countries' => 'GB',
'modified' => '2020-06-08 15:17:58',
'created' => '2020-06-08 15:17:58',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '176',
'name' => 'MicroChIP DiaPure columns SDS US en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-US-en-1_0.pdf',
'countries' => 'US',
'modified' => '2020-06-08 15:19:24',
'created' => '2020-06-08 15:19:24',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '170',
'name' => 'MicroChIP DiaPure columns SDS BE nl',
'language' => 'nl',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-nl-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:13:47',
'created' => '2020-06-08 15:13:47',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '169',
'name' => 'MicroChIP DiaPure columns SDS BE fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-fr-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:12:51',
'created' => '2020-06-08 15:12:51',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '173',
'name' => 'MicroChIP DiaPure columns SDS FR fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-FR-fr-1_0.pdf',
'countries' => 'FR',
'modified' => '2020-06-08 15:16:54',
'created' => '2020-06-08 15:16:54',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '172',
'name' => 'MicroChIP DiaPure columns SDS ES es',
'language' => 'es',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-ES-es-1_0.pdf',
'countries' => 'ES',
'modified' => '2020-06-08 15:16:02',
'created' => '2020-06-08 15:16:02',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '171',
'name' => 'MicroChIP DiaPure columns SDS DE de',
'language' => 'de',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-DE-de-1_0.pdf',
'countries' => 'DE',
'modified' => '2020-06-08 15:14:45',
'created' => '2020-06-08 15:14:45',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
)
)
)
$country = 'US'
$countries_allowed = array(
(int) 0 => 'CA',
(int) 1 => 'US',
(int) 2 => 'IE',
(int) 3 => 'GB',
(int) 4 => 'DK',
(int) 5 => 'NO',
(int) 6 => 'SE',
(int) 7 => 'FI',
(int) 8 => 'NL',
(int) 9 => 'BE',
(int) 10 => 'LU',
(int) 11 => 'FR',
(int) 12 => 'DE',
(int) 13 => 'CH',
(int) 14 => 'AT',
(int) 15 => 'ES',
(int) 16 => 'IT',
(int) 17 => 'PT'
)
$outsource = false
$other_formats = array()
$edit = ''
$testimonials = ''
$featured_testimonials = ''
$related_products = '<li>
<div class="row">
<div class="small-12 columns">
<a href="/en/p/true-microchip-kit-x16-16-rxns"><img src="/img/product/kits/chip-kit-icon.png" alt="ChIP kit icon" class="th"/></a> </div>
<div class="small-12 columns">
<div class="small-6 columns" style="padding-left:0px;padding-right:0px;margin-top:-6px;margin-left:-1px">
<span class="success label" style="">C01010132</span>
</div>
<div class="small-6 columns text-right" style="padding-left:0px;padding-right:0px;margin-top:-6px">
<!--a href="#" style="color:#B21329"><i class="fa fa-info-circle"></i></a-->
<!-- BEGIN: ADD TO CART MODAL --><div id="cartModal-1856" class="reveal-modal small" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<form action="/en/carts/add/1856" id="CartAdd/1856Form" method="post" accept-charset="utf-8"><div style="display:none;"><input type="hidden" name="_method" value="POST"/></div><input type="hidden" name="data[Cart][product_id]" value="1856" id="CartProductId"/>
<div class="row">
<div class="small-12 medium-12 large-12 columns">
<p>Add <input name="data[Cart][quantity]" placeholder="1" value="1" min="1" style="width:60px;display:inline" type="number" id="CartQuantity" required="required"/> <strong> True MicroChIP-seq Kit</strong> to my shopping cart.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="checkout" id="checkout" value="checkout" type="submit">Checkout</button> </div>
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="keepshop" id="keepshop" type="submit">Keep shopping</button> </div>
</div>
</div>
</div>
</form><a class="close-reveal-modal" aria-label="Close">×</a></div><!-- END: ADD TO CART MODAL --><a href="#" id="true-microchip-kit-x16-16-rxns" data-reveal-id="cartModal-1856" class="" style="color:#B21329"><i class="fa fa-cart-plus"></i></a>
</div>
</div>
<div class="small-12 columns" >
<h6 style="height:60px">True MicroChIP-seq Kit</h6>
</div>
</div>
</li>
'
$related = array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
'id' => '1743',
'product_id' => '1923',
'related_id' => '1856'
),
'Image' => array(
(int) 0 => array(
'id' => '1775',
'name' => 'product/kits/chip-kit-icon.png',
'alt' => 'ChIP kit icon',
'modified' => '2018-04-17 11:52:29',
'created' => '2018-03-15 15:50:34',
'ProductsImage' => array(
[maximum depth reached]
)
)
)
)
$rrbs_service = array(
(int) 0 => (int) 1894,
(int) 1 => (int) 1895
)
$chipseq_service = array(
(int) 0 => (int) 2683,
(int) 1 => (int) 1835,
(int) 2 => (int) 1836,
(int) 3 => (int) 2684,
(int) 4 => (int) 1838,
(int) 5 => (int) 1839,
(int) 6 => (int) 1856
)
$labelize = object(Closure) {
}
$old_catalog_number = ''
$country_code = 'US'
$label = '<img src="/img/banners/banner-customizer-back.png" alt=""/>'
$document = array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
'id' => '1227',
'product_id' => '1923',
'document_id' => '221'
)
)
$sds = array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
'id' => '343',
'product_id' => '1923',
'safety_sheet_id' => '175'
)
)
$publication = array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
'id' => '1173',
'product_id' => '1923',
'publication_id' => '2895'
)
)
$externalLink = ' <a href="http://www.ncbi.nlm.nih.gov/pubmed/26372092" target="_blank"><i class="fa fa-external-link"></i></a>'include - APP/View/Products/view.ctp, line 755
View::_evaluate() - CORE/Cake/View/View.php, line 971
View::_render() - CORE/Cake/View/View.php, line 933
View::render() - CORE/Cake/View/View.php, line 473
Controller::render() - CORE/Cake/Controller/Controller.php, line 963
ProductsController::slug() - APP/Controller/ProductsController.php, line 1052
ReflectionMethod::invokeArgs() - [internal], line ??
Controller::invokeAction() - CORE/Cake/Controller/Controller.php, line 491
Dispatcher::_invoke() - CORE/Cake/Routing/Dispatcher.php, line 193
Dispatcher::dispatch() - CORE/Cake/Routing/Dispatcher.php, line 167
[main] - APP/webroot/index.php, line 118
Notice (8): Undefined variable: campaign_id [APP/View/Products/view.ctp, line 755]Code Context<!-- BEGIN: REQUEST_FORM MODAL -->
<div id="request_formModal" class="reveal-modal medium" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<?= $this->element('Forms/simple_form', array('solution_of_interest' => $solution_of_interest, 'header' => $header, 'message' => $message, 'campaign_id' => $campaign_id)) ?>
$viewFile = '/home/website-server/www/app/View/Products/view.ctp'
$dataForView = array(
'language' => 'en',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'meta_title' => 'MicroChIP DiaPure columns',
'product' => array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Application' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
)
),
'Category' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Document' => array(
(int) 0 => array(
[maximum depth reached]
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
),
(int) 8 => array(
[maximum depth reached]
),
(int) 9 => array(
[maximum depth reached]
),
(int) 10 => array(
[maximum depth reached]
),
(int) 11 => array(
[maximum depth reached]
),
(int) 12 => array(
[maximum depth reached]
),
(int) 13 => array(
[maximum depth reached]
),
(int) 14 => array(
[maximum depth reached]
),
(int) 15 => array(
[maximum depth reached]
),
(int) 16 => array(
[maximum depth reached]
),
(int) 17 => array(
[maximum depth reached]
),
(int) 18 => array(
[maximum depth reached]
),
(int) 19 => array(
[maximum depth reached]
),
(int) 20 => array(
[maximum depth reached]
),
(int) 21 => array(
[maximum depth reached]
),
(int) 22 => array(
[maximum depth reached]
),
(int) 23 => array(
[maximum depth reached]
),
(int) 24 => array(
[maximum depth reached]
),
(int) 25 => array(
[maximum depth reached]
),
(int) 26 => array(
[maximum depth reached]
),
(int) 27 => array(
[maximum depth reached]
),
(int) 28 => array(
[maximum depth reached]
),
(int) 29 => array(
[maximum depth reached]
),
(int) 30 => array(
[maximum depth reached]
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
[maximum depth reached]
),
(int) 1 => array(
[maximum depth reached]
),
(int) 2 => array(
[maximum depth reached]
),
(int) 3 => array(
[maximum depth reached]
),
(int) 4 => array(
[maximum depth reached]
),
(int) 5 => array(
[maximum depth reached]
),
(int) 6 => array(
[maximum depth reached]
),
(int) 7 => array(
[maximum depth reached]
)
)
)
)
$language = 'en'
$meta_keywords = ''
$meta_description = 'MicroChIP DiaPure columns'
$meta_title = 'MicroChIP DiaPure columns'
$product = array(
'Product' => array(
'id' => '1923',
'antibody_id' => null,
'name' => 'MicroChIP DiaPure columns',
'description' => '<p><a href="https://www.diagenode.com/files/products/reagents/MicroChIP_DiaPure_manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>Diagenode’s MicroChIP DiaPure columns have been optimized for the purification and elution of very low amounts of DNA. This rapid method has been validated for epigenetic applications like low input ChIP (e.g. using the True MicroChIP kit) and CUT&Tag (e.g. using Diagenode’s pA-Tn5), but is also compatible with many other applications. The DNA can be eluted at high concentrations in volumes down to 6 μl and it is suitable for any downstream application (e.g. NGS).</p>
<p>Benefits of the MicroChIP DiaPure columns:</p>
<ul>
<li>Optimized for the purification of very low DNA amounts</li>
<li>Fast and easy protocol</li>
<li>Non-toxic</li>
<li>Validated for ChIP and Cut&Tag</li>
</ul>',
'label1' => 'Examples of results',
'info1' => '<h2 style="text-align: center;">MicroChIP DiaPure columns after ChIP</h2>
<p>Successful ChIP-seq results generated on 50,000 of K562 cells using True MicroChIP technology. ChIP has been performed accordingly to True MicroChIP protocol (Diagenode, Cat. No. C01010130), including DNA purification using the MicroChIP DiaPure columns. For the library preparation the MicroPlex Library Preparation Kit (Diagenode, Cat. No. C05010001) has been used. The below figure shows the peaks from ChIP-seq experiments using the following Diagenode antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-igv-microchip.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 1:</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50,000 of K562 cells.</p>
<p></p>
<h2 style="text-align: center;"></h2>
<h2 style="text-align: center;">MicroChIP DiaPure columns after CUT&Tag</h2>
<p>Successful CUT&Tag results showing a low background with high region-specific enrichment has been generated using 50.000 of K562 cells, 1 µg of H3K27me3 antibody (Diagenode, Cat. No. C15410069) and proteinA-Tn5 (1:250) (Diagenode, Cat. No. C01070001). 1 µg of IgG (Diagenode, Cat. No. C15410206) was used as negative control. Samples were purified using the MicroChIP DiaPure columns or phenol-chloroform purification. The below figure presenst the comparison of two purification methods.</p>
<p><img src="https://www.diagenode.com/img/product/kits/figure-diapure-igv.png" style="display: block; margin-left: auto; margin-right: auto;" /></p>
<p><strong>Figure 2:</strong> Integrative genomics viewer (IGV) visualization of CUT&Tag experiments: MicroChIP DiaPure columns vs phenol-chloroform purification using the H3K27me3 antibody.</p>',
'label2' => '',
'info2' => '',
'label3' => '',
'info3' => '',
'format' => '50 rxns',
'catalog_number' => 'C03040001',
'old_catalog_number' => '',
'sf_code' => 'C03040001-',
'type' => 'REF',
'search_order' => '04-undefined',
'price_EUR' => '130',
'price_USD' => '130',
'price_GBP' => '120',
'price_JPY' => '21300',
'price_CNY' => '',
'price_AUD' => '325',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => false,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'microchip-diapure-columns-50-rxns',
'meta_title' => 'MicroChIP DiaPure columns',
'meta_keywords' => '',
'meta_description' => 'MicroChIP DiaPure columns',
'modified' => '2023-04-20 16:11:26',
'created' => '2015-06-29 14:08:20',
'locale' => 'eng'
),
'Antibody' => array(
'host' => '*****',
'id' => null,
'name' => null,
'description' => null,
'clonality' => null,
'isotype' => null,
'lot' => null,
'concentration' => null,
'reactivity' => null,
'type' => null,
'purity' => null,
'classification' => null,
'application_table' => null,
'storage_conditions' => null,
'storage_buffer' => null,
'precautions' => null,
'uniprot_acc' => null,
'slug' => null,
'meta_keywords' => null,
'meta_description' => null,
'modified' => null,
'created' => null,
'select_label' => null
),
'Slave' => array(),
'Group' => array(),
'Related' => array(
(int) 0 => array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
[maximum depth reached]
),
'Image' => array(
[maximum depth reached]
)
)
),
'Application' => array(
(int) 0 => array(
'id' => '10',
'position' => '10',
'parent_id' => '2',
'name' => 'ChIP-qPCR',
'description' => '<div class="row">
<div class="small-12 medium-12 large-12 columns text-justify">
<p class="text-justify">Chromatin Immunoprecipitation (ChIP) coupled with quantitative PCR can be used to investigate protein-DNA interaction at known genomic binding sites. if sites are not known, qPCR primers can also be designed against potential regulatory regions such as promoters. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of performing real-time PCR is minimal. This technique is now used in a variety of life science disciplines including cellular differentiation, tumor suppressor gene silencing, and the effect of histone modifications on gene expression.</p>
<p class="text-justify"><strong>The ChIP-qPCR workflow</strong></p>
</div>
<div class="small-12 medium-12 large-12 columns text-center"><br /> <img src="https://www.diagenode.com/img/chip-qpcr-diagram.png" /></div>
<div class="small-12 medium-12 large-12 columns"><br />
<ol>
<li class="large-12 columns"><strong>Chromatin preparation: </strong>cell fixation (cross-linking) of chromatin-bound proteins such as histones or transcription factors to DNA followed by cell lysis.</li>
<li class="large-12 columns"><strong>Chromatin shearing: </strong>fragmentation of chromatin<strong> </strong>by sonication down to desired fragment size (100-500 bp)</li>
<li class="large-12 columns"><strong>Chromatin IP</strong>: protein-DNA complexe capture using<strong> <a href="https://www.diagenode.com/en/categories/chip-grade-antibodies">specific ChIP-grade antibodies</a></strong> against the histone or transcription factor of interest</li>
<li class="large-12 columns"><strong>DNA purification</strong>: chromatin reverse cross-linking and elution followed by purification<strong> </strong></li>
<li class="large-12 columns"><strong>qPCR and analysis</strong>: using previously designed primers to amplify IP'd material at specific loci</li>
</ol>
</div>
</div>
<div class="row" style="margin-top: 32px;">
<div class="small-12 medium-10 large-9 small-centered columns">
<div class="radius panel" style="background-color: #fff;">
<h3 class="text-center" style="color: #b21329;">Need guidance?</h3>
<p class="text-justify">Choose our full ChIP kits or simply choose what you need from antibodies, buffers, beads, chromatin shearing and purification reagents. With the ChIP Kit Customizer, you have complete flexibility on which components you want from our validated ChIP kits.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/which-kit-to-choose"><img src="https://www.diagenode.com/img/banners/banner-decide.png" alt="" /></a></div>
<div class="small-6 medium-6 large-6 columns"><a href="https://www.diagenode.com/pages/chip-kit-customizer-1"><img src="https://www.diagenode.com/img/banners/banner-customizer.png" alt="" /></a></div>
</div>
</div>
</div>
</div>',
'in_footer' => false,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'chip-qpcr',
'meta_keywords' => 'Chromatin immunoprecipitation,ChIP Quantitative PCR,polymerase chain reaction (PCR)',
'meta_description' => 'Diagenode's ChIP qPCR kits can be used to quantify enriched DNA after chromatin immunoprecipitation. ChIP-qPCR is advantageous in studies that focus on specific genes and potential regulatory regions across differing experimental conditions as the cost of',
'meta_title' => 'ChIP Quantitative PCR (ChIP-qPCR) | Diagenode',
'modified' => '2018-01-09 16:46:56',
'created' => '2014-12-11 00:22:08',
'ProductsApplication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '2',
'position' => '10',
'parent_id' => null,
'name' => 'エピジェネティクス・クロマチン解析',
'description' => '<div class="row">
<div class="large-12 columns">エピジェネティクス研究は、異なる転写パターン、遺伝子発現およびサイレンシングを引き起こすクロマチンの変化に対処します。<br /><br />クロマチンの主成分はDNA<span>およびヒストン蛋白質です。<span> </span></span>各ヒストンコア蛋白質(H2A<span>、</span>H2B<span>、</span>H3<span>および</span>H4<span>)の</span>2<span>つのコピーを</span>8<span>量体に組み込み、</span>DNA<span>で包んでヌクレオソームコアを形成させます。<span> </span></span>ヌクレオソームは、転写機械のDNA<span>への接近可能性および</span>クロマチン再構成因子を制御します。</div>
<div class="large-12 columns">
<p></p>
<p>クロマチン免疫沈降(ChIP<span>)は、関心対象の特定の蛋白質に対するゲノム結合部位の位置を解明するために使用される方法であり、遺伝子発現の制御に関する非常に貴重な洞察を提供します。<span> </span></span>ChIPは特定の抗原を含むクロマチン断片の選択的富化に関与します。 特定の蛋白質または蛋白質修飾を認識する抗体を使用して、特定の遺伝子座における抗原の相対存在量を決定します。</p>
<p>ChIP-seq<span>および</span>ChIP-qPCR<span>は、蛋白質</span>-DNA<span>結合部位の同定を可能にする技術です。</span></p>
<p> </p>
</div>
</div>',
'in_footer' => true,
'in_menu' => true,
'online' => true,
'tabular' => true,
'slug' => 'epigenetics-chromatin-study',
'meta_keywords' => 'Chromatin Immunoprecipitation Sequencing,ChIP-Seq,ChIP-seq grade antibodies,DNA purification,qPCR,Shearing of chromatin',
'meta_description' => 'Diagenodeは、ChIP-Seqおよびクロマチン免疫沈降(ChIP)専用キットと抗体を定量PCRアプリケーションと組み合わせて幅広く提供しています。',
'meta_title' => 'エピジェネティクス - クロマチン免疫沈降(ChIP)| Diagenode',
'modified' => '2018-01-11 09:58:47',
'created' => '2015-02-20 16:10:25',
'ProductsApplication' => array(
[maximum depth reached]
)
)
),
'Category' => array(
(int) 0 => array(
'id' => '61',
'position' => '2',
'parent_id' => '14',
'name' => 'Column-based DNA purification',
'description' => '',
'no_promo' => false,
'in_menu' => false,
'online' => true,
'tabular' => true,
'hide' => false,
'all_format' => false,
'is_antibody' => false,
'slug' => 'column-based-dna-purification',
'cookies_tag_id' => null,
'meta_keywords' => 'MicroChIP DiaPure columns,diagenode',
'meta_description' => 'Diagenode provides Column-based DNA purification for downstream analysis.',
'meta_title' => 'Column-based DNA purification | Diagenode',
'modified' => '2016-01-21 13:49:21',
'created' => '2015-07-08 10:31:04',
'ProductsCategory' => array(
[maximum depth reached]
),
'CookiesTag' => array([maximum depth reached])
)
),
'Document' => array(
(int) 0 => array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
[maximum depth reached]
)
)
),
'Feature' => array(),
'Image' => array(),
'Promotion' => array(),
'Protocol' => array(),
'Publication' => array(
(int) 0 => array(
'id' => '5033',
'name' => 'Trithorax regulates long-term memory in Drosophila through epigenetic maintenance of mushroom body metabolic state and translation capacity',
'authors' => 'Nicholas Raun et al.',
'description' => '<p><span>The role of epigenetics and chromatin in the maintenance of postmitotic neuronal cell identities is not well understood. Here, we show that the histone methyltransferase Trithorax (Trx) is required in postmitotic memory neurons of the </span><em>Drosophila</em><span><span> </span>mushroom body (MB) to enable their capacity for long-term memory (LTM), but not short-term memory (STM). Using MB-specific RNA-, ChIP-, and ATAC-sequencing, we find that Trx maintains homeostatic expression of several non-canonical MB-enriched transcripts, including the orphan nuclear receptor<span> </span></span><em>Hr51</em><span>, and the metabolic enzyme<span> </span></span><em>lactate dehydrogenase</em><span><span> </span>(</span><em>Ldh)</em><span>. Through these key targets, Trx facilitates a metabolic state characterized by high lactate levels in MBγ neurons. This metabolic state supports a high capacity for protein translation, a process that is essential for LTM, but not STM. These data suggest that Trx, a classic regulator of cell type specification during development, has additional functions in maintaining underappreciated aspects of postmitotic neuron identity, such as metabolic state. Our work supports a body of evidence suggesting that a high capacity for energy metabolism is an essential cell identity characteristic for neurons that mediate LTM.</span></p>',
'date' => '2025-01-27',
'pmid' => 'https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.3003004',
'doi' => 'https://doi.org/10.1371/journal.pbio.3003004',
'modified' => '2025-01-30 10:12:42',
'created' => '2025-01-30 10:12:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '4983',
'name' => 'Integrated multi-omics analysis of PBX1 in mouse adult neural stem- and progenitor cells identifies a transcriptional module that functionally links PBX1 to TCF3/4',
'authors' => 'Vera Laub et al.',
'description' => '<p><span>Developmental transcription factors act in networks, but how these networks achieve cell- and tissue specificity is still poorly understood. Here, we explored pre-B cell leukemia homeobox 1 (PBX1) in adult neurogenesis combining genomic, transcriptomic, and proteomic approaches. ChIP-seq analysis uncovered PBX1 binding to numerous genomic sites. Integration of PBX1 ChIP-seq with ATAC-seq data predicted interaction partners, which were subsequently validated by mass spectrometry. Whole transcriptome spatial RNA analysis revealed shared expression dynamics of </span><em>Pbx1</em><span><span> </span>and interacting factors. Among these were class I bHLH proteins TCF3 and TCF4. RNA-seq following<span> </span></span><em>Pbx1</em><span>,<span> </span></span><em>Tcf3</em><span><span> </span>or<span> </span></span><em>Tcf4</em><span><span> </span>knockdown identified proliferation- and differentiation associated genes as shared targets, while sphere formation assays following knockdown argued for functional cooperativity of PBX1 and TCF3 in progenitor cell proliferation. Notably, while physiological PBX1-TCF interaction has not yet been described, chromosomal translocation resulting in genomic<span> </span></span><em>TCF3::PBX1</em><span><span> </span>fusion characterizes a subtype of acute lymphoblastic leukemia. Introducing<span> </span></span><em>Pbx1</em><span><span> </span>into Nalm6 cells, a pre-B cell line expressing<span> </span></span><em>TCF3</em><span><span> </span>but lacking<span> </span></span><em>PBX1</em><span>, upregulated the leukemogenic genes<span> </span></span><em>BLK</em><span><span> </span>and<span> </span></span><em>NOTCH3</em><span>, arguing that functional PBX1-TCF cooperativity likely extends to hematopoiesis. Our study hence uncovers a transcriptional module orchestrating the balance between progenitor cell proliferation and differentiation in adult neurogenesis with potential implications for leukemia etiology.</span></p>',
'date' => '2024-10-08',
'pmid' => 'https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkae864/7815639',
'doi' => 'https://doi.org/10.1093/nar/gkae864',
'modified' => '2024-10-11 10:02:42',
'created' => '2024-10-11 10:02:42',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '4964',
'name' => 'A PRE loop at the dac locus acts as a topological chromatin structure that restricts and specifies enhancer–promoter communication',
'authors' => 'Denaud S. et al.',
'description' => '<p><span>Three-dimensional (3D) genome folding has a fundamental role in the regulation of developmental genes by facilitating or constraining chromatin interactions between </span><i>cis</i><span>-regulatory elements (CREs). Polycomb response elements (PREs) are a specific kind of CRE involved in the memory of transcriptional states in<span> </span></span><i>Drosophila melanogaster</i><span>. PREs act as nucleation sites for Polycomb group (PcG) proteins, which deposit the repressive histone mark H3K27me3, leading to the formation of a class of topologically associating domain (TAD) called a Polycomb domain. PREs can establish looping contacts that stabilize the gene repression of key developmental genes during development. However, the mechanism by which PRE loops fine-tune gene expression is unknown. Using clustered regularly interspaced short palindromic repeats and Cas9 genome engineering, we specifically perturbed PRE contacts or enhancer function and used complementary approaches including 4C-seq, Hi-C and Hi-M to analyze how chromatin architecture perturbation affects gene expression. Our results suggest that the PRE loop at the<span> </span></span><i>dac</i><span><span> </span>gene locus acts as a constitutive 3D chromatin scaffold during<span> </span></span><i>Drosophila</i><span><span> </span>development that forms independently of gene expression states and has a versatile function; it restricts enhancer–promoter communication and contributes to enhancer specificity.</span></p>',
'date' => '2024-08-16',
'pmid' => 'https://www.nature.com/articles/s41594-024-01375-7',
'doi' => 'https://doi.org/10.1038/s41594-024-01375-7',
'modified' => '2024-09-02 10:20:28',
'created' => '2024-09-02 10:20:28',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '4954',
'name' => 'A multiomic atlas of the aging hippocampus reveals molecular changes in response to environmental enrichment',
'authors' => 'Perez R. F. at al. ',
'description' => '<p><span>Aging involves the deterioration of organismal function, leading to the emergence of multiple pathologies. Environmental stimuli, including lifestyle, can influence the trajectory of this process and may be used as tools in the pursuit of healthy aging. To evaluate the role of epigenetic mechanisms in this context, we have generated bulk tissue and single cell multi-omic maps of the male mouse dorsal hippocampus in young and old animals exposed to environmental stimulation in the form of enriched environments. We present a molecular atlas of the aging process, highlighting two distinct axes, related to inflammation and to the dysregulation of mRNA metabolism, at the functional RNA and protein level. Additionally, we report the alteration of heterochromatin domains, including the loss of bivalent chromatin and the uncovering of a heterochromatin-switch phenomenon whereby constitutive heterochromatin loss is partially mitigated through gains in facultative heterochromatin. Notably, we observed the multi-omic reversal of a great number of aging-associated alterations in the context of environmental enrichment, which was particularly linked to glial and oligodendrocyte pathways. In conclusion, our work describes the epigenomic landscape of environmental stimulation in the context of aging and reveals how lifestyle intervention can lead to the multi-layered reversal of aging-associated decline.</span></p>',
'date' => '2024-07-16',
'pmid' => 'https://www.nature.com/articles/s41467-024-49608-z',
'doi' => 'https://doi.org/10.1038/s41467-024-49608-z',
'modified' => '2024-07-29 11:33:49',
'created' => '2024-07-29 11:33:49',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '4927',
'name' => 'Inflammatory stress-mediated chromatin changes underlie dysfunction in endothelial cells',
'authors' => 'Liu H. et al.',
'description' => '<p><span>Inflammatory stresses underlie endothelial dysfunction and contribute to the development of chronic cardiovascular disorders such as atherosclerosis and vascular fibrosis. The initial transcriptional response of endothelial cells to pro-inflammatory cytokines such as TNF-alpha is well established. However, very few studies uncover the effects of inflammatory stresses on chromatin architecture. We used integrative analysis of ATAC-seq and RNA-seq data to investigate chromatin alterations in human endothelial cells in response to TNF-alpha and febrile-range heat stress exposure. Multi-omics data analysis suggests a correlation between the transcription of stress-related genes and endothelial dysfunction drivers with chromatin regions exhibiting differential accessibility. Moreover, microscopy identified the dynamics in the nuclear organization, specifically, the changes in a subset of heterochromatic nucleoli-associated chromatin domains, the centromeres. Upon inflammatory stress exposure, the centromeres decreased association with nucleoli in a p38-dependent manner and increased the number of transcripts from pericentromeric regions. Overall, we provide two lines of evidence that suggest chromatin alterations in vascular endothelial cells during inflammatory stresses.</span></p>',
'date' => '2023-10-16',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10614786/',
'doi' => '10.1101/2023.10.11.561959',
'modified' => '2024-03-27 15:15:21',
'created' => '2024-03-27 15:15:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '4862',
'name' => 'Mutant FUS induces chromatin reorganization in the hippocampus andalters memory processes.',
'authors' => 'Tzeplaeff L. et al.',
'description' => '<p>Cytoplasmic mislocalization of the nuclear Fused in Sarcoma (FUS) protein is associated to amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Cytoplasmic FUS accumulation is recapitulated in the frontal cortex and spinal cord of heterozygous Fus mice. Yet, the mechanisms linking FUS mislocalization to hippocampal function and memory formation are still not characterized. Herein, we show that in these mice, the hippocampus paradoxically displays nuclear FUS accumulation. Multi-omic analyses showed that FUS binds to a set of genes characterized by the presence of an ETS/ELK-binding motifs, and involved in RNA metabolism, transcription, ribosome/mitochondria and chromatin organization. Importantly, hippocampal nuclei showed a decompaction of the neuronal chromatin at highly expressed genes and an inappropriate transcriptomic response was observed after spatial training of Fus mice. Furthermore, these mice lacked precision in a hippocampal-dependent spatial memory task and displayed decreased dendritic spine density. These studies shows that mutated FUS affects epigenetic regulation of the chromatin landscape in hippocampal neurons, which could participate in FTD/ALS pathogenic events. These data call for further investigation in the neurological phenotype of FUS-related diseases and open therapeutic strategies towards epigenetic drugs.</p>',
'date' => '2023-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/37327984',
'doi' => '10.1016/j.pneurobio.2023.102483',
'modified' => '2023-08-01 14:55:49',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '4733',
'name' => 'FXR inhibition may protect from SARS-CoV-2 infection by reducingACE2.',
'authors' => 'Brevini Teresa et al.',
'description' => '<p>Prevention of SARS-CoV-2 infection through the modulation of viral host receptors, such as ACE2, could represent a new chemoprophylactic approach for COVID-19 complementing vaccination. However, the mechanisms controlling ACE2 expression remain elusive. Here, we identify the farnesoid X receptor (FXR) as a direct regulator of ACE2 transcription in multiple COVID19-affected tissues, including the gastrointestinal and respiratory systems. We then use the over-the-counter compound z-guggulsterone (ZGG) and the off-patent drug ursodeoxycholic acid (UDCA) to reduce FXR signalling and downregulate ACE2 in human lung, cholangiocyte and intestinal organoids and in the corresponding tissues in mice and hamsters. We demonstrate that UDCA-mediated ACE2 downregulation reduces susceptibility to SARS-CoV-2 infection in vitro, in vivo and in human lungs and livers perfused ex situ. Furthermore, we illustrate that UDCA reduces ACE2 expression in the nasal epithelium in humans. Finally, we identify a correlation between UDCA treatment and positive clinical outcomes following SARS-CoV-2 infection using retrospective registry data, and confirm these findings in an independent validation cohort of liver transplant recipients. In conclusion, we identify a novel function of FXR in controlling ACE2 expression and provide evidence that modulation of this pathway could be beneficial for reducing SARS-CoV-2 infection, paving the road for future clinical trials.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36470304',
'doi' => '10.1038/s41586-022-05594-0',
'modified' => '2023-03-13 08:52:11',
'created' => '2023-02-28 12:19:11',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '4575',
'name' => 'Intranasal administration of Acinetobacter lwoffii in a murine model ofasthma induces IL-6-mediated protection associated with cecal microbiotachanges.',
'authors' => 'Alashkar A. B. et al.',
'description' => '<p>BACKGROUND: Early-life exposure to certain environmental bacteria including Acinetobacter lwoffii (AL) has been implicated in protection from chronic inflammatory diseases including asthma later in life. However, the underlying mechanisms at the immune-microbe interface remain largely unknown. METHODS: The effects of repeated intranasal AL exposure on local and systemic innate immune responses were investigated in wild-type and Il6 , Il10 , and Il17 mice exposed to ovalbumin-induced allergic airway inflammation. Those investigations were expanded by microbiome analyses. To assess for AL-associated changes in gene expression, the picture arising from animal data was supplemented by in vitro experiments of macrophage and T-cell responses, yielding expression and epigenetic data. RESULTS: The asthma preventive effect of AL was confirmed in the lung. Repeated intranasal AL administration triggered a proinflammatory immune response particularly characterized by elevated levels of IL-6, and consequently, IL-6 induced IL-10 production in CD4 T-cells. Both IL-6 and IL-10, but not IL-17, were required for asthma protection. AL had a profound impact on the gene regulatory landscape of CD4 T-cells which could be largely recapitulated by recombinant IL-6. AL administration also induced marked changes in the gastrointestinal microbiome but not in the lung microbiome. By comparing the effects on the microbiota according to mouse genotype and AL-treatment status, we have identified microbial taxa that were associated with either disease protection or activity. CONCLUSION: These experiments provide a novel mechanism of Acinetobacter lwoffii-induced asthma protection operating through IL-6-mediated epigenetic activation of IL-10 production and with associated effects on the intestinal microbiome.</p>',
'date' => '2022-12-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/36458896',
'doi' => '10.1111/all.15606',
'modified' => '2023-04-11 10:23:07',
'created' => '2023-02-21 09:59:46',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 8 => array(
'id' => '4836',
'name' => 'Caffeine intake exerts dual genome-wide effects on hippocampal metabolismand learning-dependent transcription.',
'authors' => 'Paiva I. et al.',
'description' => '<p>Caffeine is the most widely consumed psychoactive substance in the world. Strikingly, the molecular pathways engaged by its regular consumption remain unclear. We herein addressed the mechanisms associated with habitual (chronic) caffeine consumption in the mouse hippocampus using untargeted orthogonal omics techniques. Our results revealed that chronic caffeine exerts concerted pleiotropic effects in the hippocampus at the epigenomic, proteomic, and metabolomic levels. Caffeine lowered metabolism-related processes (e.g., at the level of metabolomics and gene expression) in bulk tissue, while it induced neuron-specific epigenetic changes at synaptic transmission/plasticity-related genes and increased experience-driven transcriptional activity. Altogether, these findings suggest that regular caffeine intake improves the signal-to-noise ratio during information encoding, in part through fine-tuning of metabolic genes, while boosting the salience of information processing during learning in neuronal circuits.</p>',
'date' => '2022-06-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35536645',
'doi' => '10.1172/JCI149371',
'modified' => '2023-08-01 13:52:29',
'created' => '2023-08-01 15:59:38',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 9 => array(
'id' => '4220',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in Mouse Prostate Cancer Xenografts',
'authors' => 'Sanchez A. et al.',
'description' => '<p><strong class="sub-title">Background/aim:<span> </span></strong>Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo.</p>
<p><strong class="sub-title">Materials and methods:<span> </span></strong>Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR.</p>
<p><strong class="sub-title">Results:<span> </span></strong>JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression.</p>
<p><strong class="sub-title">Conclusion:<span> </span></strong>JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-05-01',
'pmid' => 'https://pubmed.ncbi.nlm.nih.gov/35430567/',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-04-21 11:54:21',
'created' => '2022-04-21 11:54:21',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 10 => array(
'id' => '4409',
'name' => 'Effects of GSK-J4 on JMJD3 Histone Demethylase in MouseProstate Cancer Xenografts.',
'authors' => 'Sanchez A. et al.',
'description' => '<p>BACKGROUND/AIM: Histone methylation status is required to control gene expression. H3K27me3 is an epigenetic tri-methylation modification to histone H3 controlled by the demethylase JMJD3. JMJD3 is dysregulated in a wide range of cancers and has been shown to control the expression of a specific growth-modulatory gene signature, making it an interesting candidate to better understand prostate tumor progression in vivo. This study aimed to identify the impact of JMJD3 inhibition by its inhibitor, GSK4, on prostate tumor growth in vivo. MATERIALS AND METHODS: Prostate cancer cell lines were implanted into Balb/c nude male mice. The effects of the selective JMJD3 inhibitor GSK-J4 on tumor growth were analyzed by bioluminescence assays and H3K27me3-regulated changes in gene expression were analyzed by ChIP-qPCR and RT-qPCR. RESULTS: JMJD3 inhibition contributed to an increase in tumor growth in androgen-independent (AR-) xenografts and a decrease in androgen-dependent (AR+). GSK-J4 treatment modulated H3K27me3 enrichment on the gene panel in DU-145-luc xenografts while it had little effect on PC3-luc and no effect on LNCaP-luc. Effects of JMJD3 inhibition affected the panel gene expression. CONCLUSION: JMJD3 has a differential effect in prostate tumor progression according to AR status. Our results suggest that JMJD3 is able to play a role independently of its demethylase function in androgen-independent prostate cancer. The effects of GSK-J4 on AR+ prostate xenografts led to a decrease in tumor growth.</p>',
'date' => '2022-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/35430567',
'doi' => '10.21873/cgp.20324',
'modified' => '2022-08-11 15:11:58',
'created' => '2022-08-11 12:14:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 11 => array(
'id' => '4233',
'name' => 'Autocrine Vitamin D-signaling switches off pro-inflammatory programsof Th1 cells',
'authors' => 'Chauss D.et al.',
'description' => '<p>The molecular mechanisms governing orderly shutdown and retraction of CD4+ T helper (Th)1 responses remain poorly understood. Here, we show that complement triggers contraction of Th1 responses by inducing intrinsic expression of the vitamin D (VitD) receptor (VDR) and the VitD-activating enzyme CYP27B1, permitting T cells to both activate and respond to VitD. VitD then initiated transition from pro-inflammatory IFN-γ + Th1 cells to suppressive IL-10+ cells. This process was primed by dynamic changes in the epigenetic landscape of CD4+ T cells, generating super-enhancers and recruiting several transcription factors, notably c-JUN, STAT3 and BACH2, which together with VDR shaped the transcriptional response to VitD. Accordingly, VitD did not induce IL-10 in cells with dysfunctional BACH2 or STAT3. Bronchoalveolar lavage fluid CD4+ T cells of COVID-19 patients were Th1-skewed and showed de-repression of genes down-regulated by VitD, either from lack of substrate (VitD deficiency) and/or abnormal regulation of this system.</p>',
'date' => '2021-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/34764490',
'doi' => '10.1038/s41590-021-01080-3',
'modified' => '2022-05-19 16:57:27',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 12 => array(
'id' => '4351',
'name' => 'Essential role of a ThPOK autoregulatory loop in the maintenance ofmature CD4 T cell identity and function.',
'authors' => 'Basu Jayati et al.',
'description' => '<p>The transcription factor ThPOK (encoded by the Zbtb7b gene) controls homeostasis and differentiation of mature helper T cells, while opposing their differentiation to CD4 intraepithelial lymphocytes (IELs) in the intestinal mucosa. Thus CD4 IEL differentiation requires ThPOK transcriptional repression via reactivation of the ThPOK transcriptional silencer element (Sil). In the present study, we describe a new autoregulatory loop whereby ThPOK binds to the Sil to maintain its own long-term expression in CD4 T cells. Disruption of this loop in vivo prevents persistent ThPOK expression, leads to genome-wide changes in chromatin accessibility and derepresses the colonic regulatory T (T) cell gene expression signature. This promotes selective differentiation of naive CD4 T cells into GITRPD-1CD25 (Triple) T cells and conversion to CD4 IELs in the gut, thereby providing dominant protection from colitis. Hence, the ThPOK autoregulatory loop represents a key mechanism to physiologically control ThPOK expression and T cell differentiation in the gut, with potential therapeutic relevance.</p>',
'date' => '2021-08-01',
'pmid' => 'https://doi.org/10.1038%2Fs41590-021-00980-8',
'doi' => '10.1038/s41590-021-00980-8',
'modified' => '2022-06-22 12:32:59',
'created' => '2022-05-19 10:41:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 13 => array(
'id' => '4127',
'name' => 'The histone modification H3K4me3 is altered at the locus in Alzheimer'sdisease brain.',
'authors' => 'Smith, Adam et al.',
'description' => '<p>Several epigenome-wide association studies of DNA methylation have highlighted altered DNA methylation in the gene in Alzheimer's disease (AD) brain samples. However, no study has specifically examined histone modifications in the disease. We use chromatin immunoprecipitation-qPCR to quantify tri-methylation at histone 3 lysine 4 (H3K4me3) and 27 (H3K27me3) in the gene in entorhinal cortex from donors with high (n = 59) or low (n = 29) Alzheimer's disease pathology. We demonstrate decreased levels of H3K4me3, a marker of active gene transcription, with no change in H3K27me3, a marker of inactive genes. H3K4me3 is negatively correlated with DNA methylation in specific regions of the gene. Our study suggests that the gene shows altered epigenetic marks indicative of reduced gene activation in Alzheimer's disease.</p>',
'date' => '2021-02-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/33815817',
'doi' => '10.2144/fsoa-2020-0161',
'modified' => '2021-12-07 10:16:08',
'created' => '2021-12-06 15:53:19',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 14 => array(
'id' => '4062',
'name' => 'Digging Deeper into Breast Cancer Epigenetics: Insights from ChemicalInhibition of Histone Acetyltransferase TIP60 .',
'authors' => 'Idrissou, Mouhamed and Lebert, Andre and Boisnier, Tiphanie and Sanchez,Anna and Houfaf Khoufaf, Fatma Zohra and Penault-Llorca, Frédérique andBignon, Yves-Jean and Bernard-Gallon, Dominique',
'description' => '<p>Breast cancer is often sporadic due to several factors. Among them, the deregulation of epigenetic proteins may be involved. TIP60 or KAT5 is an acetyltransferase that regulates gene transcription through the chromatin structure. This pleiotropic protein acts in several cellular pathways by acetylating proteins. RNA and protein expressions of TIP60 were shown to decrease in some breast cancer subtypes, particularly in triple-negative breast cancer (TNBC), where a low expression of TIP60 was exhibited compared with luminal subtypes. In this study, the inhibition of the residual activity of TIP60 in breast cancer cell lines was investigated by using two chemical inhibitors, TH1834 and NU9056, first on the acetylation of the specific target, lysine 4 of histone 3 (H3K4) by immunoblotting, and second, by chromatin immunoprecipitation (ChIP)-qPCR (-quantitative Polymerase Chain Reaction). Subsequently, significant decreases or a trend toward decrease of H3K4ac in the different chromatin compartments were observed. In addition, the expression of 48 human nuclear receptors was studied with TaqMan Low-Density Array in these breast cancer cell lines treated with TIP60 inhibitors. The statistical analysis allowed us to comprehensively characterize the androgen receptor and receptors in TNBC cell lines after TH1834 or NU9056 treatment. The understanding of the residual activity of TIP60 in the evolution of breast cancer might be a major asset in the fight against this disease, and could allow TIP60 to be used as a biomarker or therapeutic target for breast cancer progression in the future.</p>',
'date' => '2020-10-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/32960142',
'doi' => '10.1089/omi.2020.0104',
'modified' => '2021-02-19 17:39:52',
'created' => '2021-02-18 10:21:53',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 15 => array(
'id' => '3969',
'name' => 'Delineating the early transcriptional specification of the mammalian trachea and esophagus.',
'authors' => 'Kuwahara A, Lewis AE, Coombes C, Leung FS, Percharde M, Bush JO',
'description' => '<p>The genome-scale transcriptional programs that specify the mammalian trachea and esophagus are unknown. Though NKX2-1 and SOX2 are hypothesized to be co-repressive master regulators of tracheoesophageal fates, this is untested at a whole transcriptomic scale and their downstream networks remain unidentified. By combining single-cell RNA-sequencing with bulk RNA-sequencing of mutants and NKX2-1 ChIP-sequencing in mouse embryos, we delineate the NKX2-1 transcriptional program in tracheoesophageal specification, and discover that the majority of the tracheal and esophageal transcriptome is NKX2-1 independent. To decouple the NKX2-1 transcriptional program from regulation by SOX2, we interrogate the expression of newly-identified tracheal and esophageal markers in / compound mutants. Finally, we discover that NKX2-1 binds directly to and and regulates their expression to control mesenchymal specification to cartilage and smooth muscle, coupling epithelial identity with mesenchymal specification. These findings create a new framework for understanding early tracheoesophageal fate specification at the genome-wide level.</p>',
'date' => '2020-06-09',
'pmid' => 'http://www.pubmed.gov/32515350',
'doi' => '10.7554/eLife.55526',
'modified' => '2020-08-12 09:32:02',
'created' => '2020-08-10 12:12:25',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 16 => array(
'id' => '3845',
'name' => 'Combinatorial action of NF-Y and TALE at embryonic enhancers defines distinct gene expression programs during zygotic genome activation in zebrafish.',
'authors' => 'Stanney W, Ladam F, Donaldson IJ, Parsons TJ, Maehr R, Bobola N, Sagerström CG',
'description' => '<p>Animal embryogenesis is initiated by maternal factors, but zygotic genome activation (ZGA) shifts regulatory control to the embryo during blastula stages. ZGA is thought to be mediated by maternally provided transcription factors (TFs), but few such TFs have been identified in vertebrates. Here we report that NF-Y and TALE TFs bind zebrafish genomic elements associated with developmental control genes already at ZGA. In particular, co-regulation by NF-Y and TALE is associated with broadly acting genes involved in transcriptional control, while regulation by either NF-Y or TALE defines genes in specific developmental processes, such that NF-Y controls a cilia gene expression program while TALE controls expression of hox genes. We also demonstrate that NF-Y and TALE-occupied genomic elements function as enhancers during embryogenesis. We conclude that combinatorial use of NF-Y and TALE at developmental enhancers permits the establishment of distinct gene expression programs at zebrafish ZGA.</p>',
'date' => '2019-12-17',
'pmid' => 'http://www.pubmed.gov/31862379',
'doi' => '10.1016/j.ydbio.2019.12.003',
'modified' => '2020-02-20 11:13:27',
'created' => '2020-02-13 10:02:44',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 17 => array(
'id' => '3744',
'name' => 'Whsc1 links pluripotency exit with mesendoderm specification.',
'authors' => 'Tian TV, Di Stefano B, Stik G, Vila-Casadesús M, Sardina JL, Vidal E, Dasti A, Segura-Morales C, De Andrés-Aguayo L, Gómez A, Goldmann J, Jaenisch R, Graf T',
'description' => '<p>How pluripotent stem cells differentiate into the main germ layers is a key question of developmental biology. Here, we show that the chromatin-related factor Whsc1 (also known as Nsd2 and MMSET) has a dual role in pluripotency exit and germ layer specification of embryonic stem cells. On induction of differentiation, a proportion of Whsc1-depleted embryonic stem cells remain entrapped in a pluripotent state and fail to form mesendoderm, although they are still capable of generating neuroectoderm. These functions of Whsc1 are independent of its methyltransferase activity. Whsc1 binds to enhancers of the mesendodermal regulators Gata4, T (Brachyury), Gata6 and Foxa2, together with Brd4, and activates the expression of these genes. Depleting each of these regulators also delays pluripotency exit, suggesting that they mediate the effects observed with Whsc1. Our data indicate that Whsc1 links silencing of the pluripotency regulatory network with activation of mesendoderm lineages.</p>',
'date' => '2019-07-01',
'pmid' => 'http://www.pubmed.gov/31235934',
'doi' => '10.1038/s41556-019-0342-1',
'modified' => '2019-08-06 16:35:35',
'created' => '2019-07-31 13:35:50',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 18 => array(
'id' => '3639',
'name' => 'Spatial confinement downsizes the inflammatory response of macrophages.',
'authors' => 'Jain N, Vogel V',
'description' => '<p>Macrophages respond to chemical/metabolic and physical stimuli, but their effects cannot be readily decoupled in vivo during pro-inflammatory activation. Here, we show that preventing macrophage spreading by spatial confinement, as imposed by micropatterning, microporous substrates or cell crowding, suppresses late lipopolysaccharide (LPS)-activated transcriptional programs (biomarkers IL-6, CXCL9, IL-1β, and iNOS) by mechanomodulating chromatin compaction and epigenetic alterations (HDAC3 levels and H3K36-dimethylation). Mechanistically, confinement reduces actin polymerization, thereby lowers the LPS-stimulated nuclear translocation of MRTF-A. This lowers the activity of the MRTF-A-SRF complex and subsequently downregulates the inflammatory response, as confirmed by chromatin immunoprecipitation coupled with quantitative PCR and RNA sequencing analysis. Confinement thus downregulates pro-inflammatory cytokine secretion and, well before any activation processes, the phagocytic potential of macrophages. Contrarily, early events, including activation of the LPS receptor TLR4, and downstream NF-κB and IRF3 signalling and hence the expression of early LPS-responsive genes were marginally affected by confinement. These findings have broad implications in the context of mechanobiology, inflammation and immunology, as well as in tissue engineering and regenerative medicine.</p>',
'date' => '2018-12-01',
'pmid' => 'http://www.pubmed.gov/30349032',
'doi' => '10.1038/s41563-018-0190-6',
'modified' => '2019-06-07 10:23:26',
'created' => '2019-06-06 12:11:18',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 19 => array(
'id' => '3379',
'name' => 'SIRT1-dependent epigenetic regulation of H3 and H4 histone acetylation in human breast cancer',
'authors' => 'Khaldoun Rifaï et al.',
'description' => '<p>Breast cancer is the most frequently diagnosed malignancy in women worldwide. It is well established that the complexity of carcinogenesis involves profound epigenetic deregulations that contribute to the tumorigenesis process. Deregulated H3 and H4 acetylated histone marks are amongst those alterations. Sirtuin-1 (SIRT1) is a class-III histone deacetylase deeply involved in apoptosis, genomic stability, gene expression regulation and breast tumorigenesis. However, the underlying molecular mechanism by which SIRT1 regulates H3 and H4 acetylated marks, and consequently cancer-related gene expression in breast cancer, remains uncharacterized. In this study, we elucidated SIRT1 epigenetic role and analyzed the link between the latter and histones H3 and H4 epigenetic marks in all 5 molecular subtypes of breast cancer. Using a cohort of 135 human breast tumors and their matched normal tissues, as well as 5 human-derived cell lines, we identified H3k4ac as a new prime target of SIRT1 in breast cancer. We also uncovered an inverse correlation between SIRT1 and the 3 epigenetic marks H3k4ac, H3k9ac and H4k16ac expression patterns. We showed that SIRT1 modulates the acetylation patterns of histones H3 and H4 in breast cancer. Moreover, SIRT1 regulates its H3 acetylated targets in a subtype-specific manner. Furthermore, SIRT1 siRNA-mediated knockdown increases histone acetylation levels at 6 breast cancer-related gene promoters: <em>AR</em>, <em>BRCA1</em>, <em>ERS1</em>, <em>ERS2</em>, <em>EZH2</em> and <em>EP300</em>. In summary, this report characterizes for the first time the epigenetic behavior of SIRT1 in human breast carcinoma. These novel findings point to a potential use of SIRT1 as an epigenetic therapeutic target in breast cancer.</p>',
'date' => '2018-07-17',
'pmid' => 'http://www.oncotarget.com/index.php?journal=oncotarget&page=article&op=view&path[]=25771&path[]=80619',
'doi' => '',
'modified' => '2018-08-09 10:47:58',
'created' => '2018-07-26 12:02:12',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 20 => array(
'id' => '3595',
'name' => 'Reciprocal signalling by Notch-Collagen V-CALCR retains muscle stem cells in their niche.',
'authors' => 'Baghdadi MB, Castel D, Machado L, Fukada SI, Birk DE, Relaix F, Tajbakhsh S, Mourikis P',
'description' => '<p>The cell microenvironment, which is critical for stem cell maintenance, contains both cellular and non-cellular components, including secreted growth factors and the extracellular matrix. Although Notch and other signalling pathways have previously been reported to regulate quiescence of stem cells, the composition and source of molecules that maintain the stem cell niche remain largely unknown. Here we show that adult muscle satellite (stem) cells in mice produce extracellular matrix collagens to maintain quiescence in a cell-autonomous manner. Using chromatin immunoprecipitation followed by sequencing, we identified NOTCH1/RBPJ-bound regulatory elements adjacent to specific collagen genes, the expression of which is deregulated in Notch-mutant mice. Moreover, we show that Collagen V (COLV) produced by satellite cells is a critical component of the quiescent niche, as depletion of COLV by conditional deletion of the Col5a1 gene leads to anomalous cell cycle entry and gradual diminution of the stem cell pool. Notably, the interaction of COLV with satellite cells is mediated by the Calcitonin receptor, for which COLV acts as a surrogate local ligand. Systemic administration of a calcitonin derivative is sufficient to rescue the quiescence and self-renewal defects found in COLV-null satellite cells. This study reveals a Notch-COLV-Calcitonin receptor signalling cascade that maintains satellite cells in a quiescent state in a cell-autonomous fashion, and raises the possibility that similar reciprocal mechanisms act in diverse stem cell populations.</p>',
'date' => '2018-05-23',
'pmid' => 'http://www.pubmed.gov/29795344',
'doi' => '10.1038/s41586-018-0144-9',
'modified' => '2019-04-17 15:12:55',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 21 => array(
'id' => '3589',
'name' => 'A new metabolic gene signature in prostate cancer regulated by JMJD3 and EZH2.',
'authors' => 'Daures M, Idrissou M, Judes G, Rifaï K, Penault-Llorca F, Bignon YJ, Guy L, Bernard-Gallon D',
'description' => '<p>Histone methylation is essential for gene expression control. Trimethylated lysine 27 of histone 3 (H3K27me3) is controlled by the balance between the activities of JMJD3 demethylase and EZH2 methyltransferase. This epigenetic mark has been shown to be deregulated in prostate cancer, and evidence shows H3K27me3 enrichment on gene promoters in prostate cancer. To study the impact of this enrichment, a transcriptomic analysis with TaqMan Low Density Array (TLDA) of several genes was studied on prostate biopsies divided into three clinical grades: normal ( = 23) and two tumor groups that differed in their aggressiveness (Gleason score ≤ 7 ( = 20) and >7 ( = 19)). ANOVA demonstrated that expression of the gene set was upregulated in tumors and correlated with Gleason score, thus discriminating between the three clinical groups. Six genes involved in key cellular processes stood out: , , , , and . Chromatin immunoprecipitation demonstrated collocation of EZH2 and JMJD3 on gene promoters that was dependent on disease stage. Gene set expression was also evaluated on prostate cancer cell lines (DU 145, PC-3 and LNCaP) treated with an inhibitor of JMJD3 (GSK-J4) or EZH2 (DZNeP) to study their involvement in gene regulation. Results showed a difference in GSK-J4 sensitivity under PTEN status of cell lines and an opposite gene expression profile according to androgen status of cells. In summary, our data describe the impacts of JMJD3 and EZH2 on a new gene signature involved in prostate cancer that may help identify diagnostic and therapeutic targets in prostate cancer.</p>',
'date' => '2018-05-04',
'pmid' => 'http://www.pubmed.gov/29805743',
'doi' => '10.18632/oncotarget.25182',
'modified' => '2019-04-17 15:21:33',
'created' => '2019-04-16 12:25:30',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 22 => array(
'id' => '3477',
'name' => 'Contrasting epigenetic states of heterochromatin in the different types of mouse pluripotent stem cells.',
'authors' => 'Tosolini M, Brochard V, Adenot P, Chebrout M, Grillo G, Navia V, Beaujean N, Francastel C, Bonnet-Garnier A, Jouneau A',
'description' => '<p>Mouse embryonic stem cells (ESCs) and epiblast stem cells (EpiSCs) represent naive and primed pluripotency states, respectively, and are maintained in vitro by specific signalling pathways. Furthermore, ESCs cultured in serum-free medium with two kinase inhibitors (2i-ESCs) are thought to be the ground naïve pluripotent state. Here, we present a comparative study of the epigenetic and transcriptional states of pericentromeric heterochromatin satellite sequences found in these pluripotent states. We show that 2i-ESCs are distinguished from other pluripotent cells by a prominent enrichment in H3K27me3 and low levels of DNA methylation at pericentromeric heterochromatin. In contrast, serum-containing ESCs exhibit higher levels of major satellite repeat transcription, which is lower in 2i-ESCs and even more repressed in primed EpiSCs. Removal of either DNA methylation or H3K9me3 at PCH in 2i-ESCs leads to enhanced deposition of H3K27me3 with few changes in satellite transcript levels. In contrast, their removal in EpiSCs does not lead to deposition of H3K27me3 but rather removes transcriptional repression. Altogether, our data show that the epigenetic state of PCH is modified during transition from naive to primed pluripotency states towards a more repressive state, which tightly represses the transcription of satellite repeats.</p>',
'date' => '2018-04-10',
'pmid' => 'http://www.pubmed.gov/29636490',
'doi' => '10.1038/s41598-018-23822-4',
'modified' => '2019-02-15 20:26:34',
'created' => '2019-02-14 15:01:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 23 => array(
'id' => '3324',
'name' => 'Isl1 mediates mesenchymal expansion in the developing external genitalia via regulation of Bmp4, Fgf10 and Wnt5a',
'authors' => 'Ching S.T. et al.',
'description' => '<p>Genital malformations are among the most common human birth defects, and both genetic and environmental factors can contribute to these malformations. Development of the external genitalia in mammals relies on complex signaling networks, and disruption of these signaling pathways can lead to genital defects. Islet-1 (ISL1), a member of the LIM/Homeobox family of transcription factors, has been identified as a major susceptibility gene for classic bladder exstrophy in humans, a common form of the bladder exstrophy-epispadias complex (BEEC), and is implicated in a role in urinary tract development. We report that deletion of Isl1 from the genital mesenchyme in mice led to hypoplasia of the genital tubercle and prepuce, with an ectopic urethral opening and epispadias-like phenotype. These mice also developed hydroureter and hydronephrosis. Identification of ISL1 transcriptional targets via ChIP-Seq and expression analyses revealed that Isl1 regulates several important signaling pathways during embryonic genital development, including the BMP, WNT, and FGF cascades. An essential function of Isl1 during development of the external genitalia is to induce Bmp4-mediated apoptosis in the genital mesenchyme. Together, these studies demonstrate that Isl1 plays a critical role during development of the external genitalia and forms the basis for a greater understanding of the molecular mechanisms underlying the pathogenesis of BEEC and urinary tract defects in humans.</p>',
'date' => '2018-01-01',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/29126155',
'doi' => '',
'modified' => '2018-02-06 09:25:22',
'created' => '2018-02-06 09:25:22',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 24 => array(
'id' => '3322',
'name' => 'In Situ Fixation Redefines Quiescence and Early Activation of Skeletal Muscle Stem Cells',
'authors' => 'Machado L. et al.',
'description' => '<div class="abstract">
<h2 class="sectionTitle" tabindex="0">Summary</h2>
<div class="content">
<p>State of the art techniques have been developed to isolate and analyze cells from various tissues, aiming to capture their <em>in vivo</em> state. However, the majority of cell isolation protocols involve lengthy mechanical and enzymatic dissociation steps followed by flow cytometry, exposing cells to stress and disrupting their physiological niche. Focusing on adult skeletal muscle stem cells, we have developed a protocol that circumvents the impact of isolation procedures and captures cells in their native quiescent state. We show that current isolation protocols induce major transcriptional changes accompanied by specific histone modifications while having negligible effects on DNA methylation. In addition to proposing a protocol to avoid isolation-induced artifacts, our study reveals previously undetected quiescence and early activation genes of potential biological interest.</p>
</div>
</div>',
'date' => '2017-11-14',
'pmid' => 'http://www.cell.com/cell-reports/abstract/S2211-1247(17)31543-7',
'doi' => '',
'modified' => '2022-05-19 16:11:43',
'created' => '2018-02-02 16:36:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 25 => array(
'id' => '3263',
'name' => 'The RPMI-1640 vitamin mixture promotes bovine blastocyst development in vitro and downregulates gene expression of TXNIP with epigenetic modification of associated histones',
'authors' => 'Ikeda S.et al.',
'description' => '<p>Diverse environmental conditions surrounding preimplantation embryos, including available nutrients, affect their metabolism and development in both short- and long-term manner. Thioredoxin-interacting protein (TXNIP) is a possible marker for preimplantation stress that is implicated in in vitro fertilization- (IVF) induced long-term DOHaD effects. B vitamins, as participants in one-carbon metabolism, may affect preimplantation embryos by epigenetic alterations of metabolically and developmentally important genes. In vitro-produced bovine embryos were cultured with or without Roswell Park Memorial Institute 1640 vitamin mixture, containing B vitamins and B vitamin-like substances, from day 3 after IVF and we evaluated blastocyst development and TXNIP messenger RNA (mRNA) expression in the blastocysts by reverse transcription-quantitative polymerase chain reaction. The degree of trimethylation of histone H3 lysine 27 (H3K27me3) at TXNIP promoter was examined semi-quantitatively by chromatin immunoprecipitation polymerase chain reaction. Total H3K27me3 were also compared between the groups by Western blot analysis. The vitamin treatment significantly increased the rates of blastocyst development (P<0.05) and their hatching (P<0.001) from the zona pellucida by day 8. The mRNA expression of TXNIP was lower (P<0.01) in blastocysts in the vitamin-mixture-treated group concomitant with higher (P<0.05) level of H3K27me3 of its promoter compared with the control group. The total H3K27me3 in the vitamin-mixture-treated group was also higher (P<0.01) than that in the control group. The epigenetic control of genes related to important metabolic processes during the periconceptional period by nutritional conditions in utero and/or in vitro may have possible implication for the developmental programming during this period that may impact the welfare and production traits of farm animals.</p>',
'date' => '2017-08-02',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28764817',
'doi' => '',
'modified' => '2017-10-09 15:59:51',
'created' => '2017-10-09 15:59:51',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 26 => array(
'id' => '3193',
'name' => 'Global analysis of H3K27me3 as an epigenetic marker in prostate cancer progression',
'authors' => 'Ngollo M. et al.',
'description' => '<div class="">
<h4>BACKGROUND:</h4>
<p><abstracttext label="BACKGROUND" nlmcategory="BACKGROUND">H3K27me3 histone marks shape the inhibition of gene transcription. In prostate cancer, the deregulation of H3K27me3 marks might play a role in prostate tumor progression.</abstracttext></p>
<h4>METHODS:</h4>
<p><abstracttext label="METHODS" nlmcategory="METHODS">We investigated genome-wide H3K27me3 histone methylation profile using chromatin immunoprecipitation (ChIP) and 2X400K promoter microarrays to identify differentially-enriched regions in biopsy samples from prostate cancer patients. H3K27me3 marks were assessed in 34 prostate tumors: 11 with Gleason score > 7 (GS > 7), 10 with Gleason score ≤ 7 (GS ≤ 7), and 13 morphologically normal prostate samples.</abstracttext></p>
<h4>RESULTS:</h4>
<p><abstracttext label="RESULTS" nlmcategory="RESULTS">Here, H3K27me3 profiling identified an average of 386 enriched-genes on promoter regions in healthy control group versus 545 genes in GS ≤ 7 and 748 genes in GS > 7 group. We then ran a factorial discriminant analysis (FDA) and compared the enriched genes in prostate-tumor biopsies and normal biopsies using ANOVA to identify significantly differentially-enriched genes. The analysis identified ALG5, EXOSC8, CBX1, GRID2, GRIN3B, ING3, MYO1D, NPHP3-AS1, MSH6, FBXO11, SND1, SPATS2, TENM4 and TRA2A genes. These genes are possibly associated with prostate cancer. Notably, the H3K27me3 histone mark emerged as a novel regulatory mechanism in poor-prognosis prostate cancer.</abstracttext></p>
<h4>CONCLUSIONS:</h4>
<p><abstracttext label="CONCLUSIONS" nlmcategory="CONCLUSIONS">Our findings point to epigenetic mark H3K27me3 as an important event in prostate carcinogenesis and progression. The results reported here provide new molecular insights into the pathogenesis of prostate cancer.</abstracttext></p>
</div>',
'date' => '2017-04-12',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28403887',
'doi' => '',
'modified' => '2017-06-19 14:07:35',
'created' => '2017-06-19 14:05:03',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 27 => array(
'id' => '3156',
'name' => 'Crebbp loss cooperates with Bcl2 over-expression to promote lymphoma in mice',
'authors' => 'Idoia García-Ramírez, Saber Tadros, Inés González-Herrero, Alberto Martín-Lorenzo, Guillermo Rodríguez-Hernández, Dalia Moore, Lucía Ruiz-Roca, Oscar Blanco, Diego Alonso-López, Javier De Las Rivas, Keenan Hartert, Romain Duval, David Klinkebiel, Martin B',
'description' => '<p><span>CREBBP is targeted by inactivating mutations in follicular lymphoma (FL) and diffuse large B-cell lymphoma (DLBCL). Here, we provide evidence from transgenic mouse models that Crebbp deletion results in deficits in B-cell development and can cooperate with Bcl2 over-expression to promote B-cell lymphoma. Through transcriptional and epigenetic profiling of these B-cells we found that Crebbp inactivation was associated with broad transcriptional alterations, but no changes in the patterns of histone acetylation at the proximal regulatory regions of these genes. However, B-cells with Crebbp inactivation showed high expression of Myc and patterns of altered histone acetylation that were localized to intragenic regions, enriched for Myc DNA binding motifs, and showed Myc binding. Through the analysis of CREBBP mutations from a large cohort of primary human FL and DLBCL, we show a significant difference in the spectrum of CREBBP mutations in these two diseases, with higher frequencies of nonsense/frameshift mutations in DLBCL compared to FL. Together our data therefore provide important links between Crebbp inactivation and Bcl2 dependence, and show a role for Crebbp inactivation in the induction of Myc expression. We suggest this may parallel the role of CREBBP frameshift/nonsense mutations in DLBCL that result in loss of the protein, but may contrast the role of missense mutations in the lysine acetyltransferase domain that are more frequently observed in FL and yield an inactive protein.</span></p>',
'date' => '2017-03-05',
'pmid' => 'http://www.bloodjournal.org/content/early/2017/03/13/blood-2016-08-733469?sso-checked=true',
'doi' => 'https://doi.org/10.1182/blood-2016-08-733469',
'modified' => '2017-05-11 11:17:42',
'created' => '2017-04-10 07:56:37',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 28 => array(
'id' => '3151',
'name' => 'Aorta macrophage inflammatory and epigenetic changes in a murine model of obstructive sleep apnea: Potential role of CD36.',
'authors' => 'Cortese R. et al.',
'description' => '<p>Obstructive sleep apnea (OSA) affects 8-10% of the population, is characterized by chronic intermittent hypoxia (CIH), and causally associates with cardiovascular morbidities. In CIH-exposed mice, closely mimicking the chronicity of human OSA, increased accumulation and proliferation of pro-inflammatory metabolic M1-like macrophages highly expressing CD36, emerged in aorta. Transcriptomic and MeDIP-seq approaches identified activation of pro-atherogenic pathways involving a complex interplay of histone modifications in functionally-relevant biological pathways, such as inflammation and oxidative stress in aorta macrophages. Discontinuation of CIH did not elicit significant improvements in aorta wall macrophage phenotype. However, CIH-induced aorta changes were absent in CD36 knockout mice, Our results provide mechanistic insights showing that CIH exposures during sleep in absence of concurrent pro-atherogenic settings (i.e., genetic propensity or dietary manipulation) lead to the recruitment of CD36(+)<sup>high</sup> macrophages to the aortic wall and trigger atherogenesis. Furthermore, long-term CIH-induced changes may not be reversible with usual OSA treatment.</p>',
'date' => '2017-02-27',
'pmid' => 'https://www.ncbi.nlm.nih.gov/pubmed/28240319',
'doi' => '',
'modified' => '2017-03-28 09:16:02',
'created' => '2017-03-28 09:16:02',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 29 => array(
'id' => '3029',
'name' => 'Loss of Ezh2 synergizes with JAK2-V617F in initiating myeloproliferative neoplasms and promoting myelofibrosis',
'authors' => 'Shimizu T et al.',
'description' => '<p>Myeloproliferative neoplasm (MPN) patients frequently show co-occurrence of <em>JAK2</em>-V617F and mutations in epigenetic regulator genes, including <em>EZH2</em>. In this study, we show that <em>JAK2</em>-V617F and loss of <em>Ezh2</em> in hematopoietic cells contribute synergistically to the development of MPN. The MPN phenotype induced by <em>JAK2</em>-V617F was accentuated in <em>JAK2</em>-V617F<em>;Ezh2<sup>−/−</sup></em> mice, resulting in very high platelet and neutrophil counts, more advanced myelofibrosis, and reduced survival. These mice also displayed expansion of the stem cell and progenitor cell compartments and a shift of differentiation toward megakaryopoiesis at the expense of erythropoiesis. Single cell limiting dilution transplantation with bone marrow from <em>JAK2</em>-V617F<em>;Ezh2<sup>+/−</sup></em> mice showed increased reconstitution and MPN disease initiation potential compared with <em>JAK2</em>-V617F alone. RNA sequencing in <em>Ezh2</em>-deficient hematopoietic stem cells (HSCs) and megakaryocytic erythroid progenitors identified highly up-regulated genes, including <em>Lin28b</em> and <em>Hmga2</em>, and chromatin immunoprecipitation (ChIP)–quantitative PCR (qPCR) analysis of their promoters revealed decreased H3K27me3 deposition. Forced expression of <em>Hmga2</em> resulted in increased chimerism and platelet counts in recipients of retrovirally transduced HSCs. <em>JAK2</em>-V617F–expressing mice treated with an Ezh2 inhibitor showed higher platelet counts than vehicle controls. Our data support the proposed tumor suppressor function of <em>EZH2</em> in patients with MPN and call for caution when considering using Ezh2 inhibitors in MPN.</p>',
'date' => '2016-07-11',
'pmid' => 'http://jem.rupress.org/content/213/8/1479.short?rss=1',
'doi' => '',
'modified' => '2016-09-08 15:24:48',
'created' => '2016-09-08 15:24:48',
'ProductsPublication' => array(
[maximum depth reached]
)
),
(int) 30 => array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
[maximum depth reached]
)
)
),
'Testimonial' => array(),
'Area' => array(),
'SafetySheet' => array(
(int) 0 => array(
'id' => '174',
'name' => 'MicroChIP DiaPure columns SDS GB en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-GB-en-1_0.pdf',
'countries' => 'GB',
'modified' => '2020-06-08 15:17:58',
'created' => '2020-06-08 15:17:58',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 1 => array(
'id' => '176',
'name' => 'MicroChIP DiaPure columns SDS US en',
'language' => 'en',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-US-en-1_0.pdf',
'countries' => 'US',
'modified' => '2020-06-08 15:19:24',
'created' => '2020-06-08 15:19:24',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 2 => array(
'id' => '170',
'name' => 'MicroChIP DiaPure columns SDS BE nl',
'language' => 'nl',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-nl-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:13:47',
'created' => '2020-06-08 15:13:47',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 3 => array(
'id' => '169',
'name' => 'MicroChIP DiaPure columns SDS BE fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-BE-fr-1_0.pdf',
'countries' => 'BE',
'modified' => '2020-06-08 15:12:51',
'created' => '2020-06-08 15:12:51',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 4 => array(
'id' => '173',
'name' => 'MicroChIP DiaPure columns SDS FR fr',
'language' => 'fr',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-FR-fr-1_0.pdf',
'countries' => 'FR',
'modified' => '2020-06-08 15:16:54',
'created' => '2020-06-08 15:16:54',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 5 => array(
'id' => '172',
'name' => 'MicroChIP DiaPure columns SDS ES es',
'language' => 'es',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-ES-es-1_0.pdf',
'countries' => 'ES',
'modified' => '2020-06-08 15:16:02',
'created' => '2020-06-08 15:16:02',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 6 => array(
'id' => '171',
'name' => 'MicroChIP DiaPure columns SDS DE de',
'language' => 'de',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-DE-de-1_0.pdf',
'countries' => 'DE',
'modified' => '2020-06-08 15:14:45',
'created' => '2020-06-08 15:14:45',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
),
(int) 7 => array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
[maximum depth reached]
)
)
)
)
$country = 'US'
$countries_allowed = array(
(int) 0 => 'CA',
(int) 1 => 'US',
(int) 2 => 'IE',
(int) 3 => 'GB',
(int) 4 => 'DK',
(int) 5 => 'NO',
(int) 6 => 'SE',
(int) 7 => 'FI',
(int) 8 => 'NL',
(int) 9 => 'BE',
(int) 10 => 'LU',
(int) 11 => 'FR',
(int) 12 => 'DE',
(int) 13 => 'CH',
(int) 14 => 'AT',
(int) 15 => 'ES',
(int) 16 => 'IT',
(int) 17 => 'PT'
)
$outsource = false
$other_formats = array()
$edit = ''
$testimonials = ''
$featured_testimonials = ''
$related_products = '<li>
<div class="row">
<div class="small-12 columns">
<a href="/en/p/true-microchip-kit-x16-16-rxns"><img src="/img/product/kits/chip-kit-icon.png" alt="ChIP kit icon" class="th"/></a> </div>
<div class="small-12 columns">
<div class="small-6 columns" style="padding-left:0px;padding-right:0px;margin-top:-6px;margin-left:-1px">
<span class="success label" style="">C01010132</span>
</div>
<div class="small-6 columns text-right" style="padding-left:0px;padding-right:0px;margin-top:-6px">
<!--a href="#" style="color:#B21329"><i class="fa fa-info-circle"></i></a-->
<!-- BEGIN: ADD TO CART MODAL --><div id="cartModal-1856" class="reveal-modal small" data-reveal aria-labelledby="modalTitle" aria-hidden="true" role="dialog">
<form action="/en/carts/add/1856" id="CartAdd/1856Form" method="post" accept-charset="utf-8"><div style="display:none;"><input type="hidden" name="_method" value="POST"/></div><input type="hidden" name="data[Cart][product_id]" value="1856" id="CartProductId"/>
<div class="row">
<div class="small-12 medium-12 large-12 columns">
<p>Add <input name="data[Cart][quantity]" placeholder="1" value="1" min="1" style="width:60px;display:inline" type="number" id="CartQuantity" required="required"/> <strong> True MicroChIP-seq Kit</strong> to my shopping cart.</p>
<div class="row">
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="checkout" id="checkout" value="checkout" type="submit">Checkout</button> </div>
<div class="small-6 medium-6 large-6 columns">
<button class="alert small button expand" onclick="$(this).addToCart('True MicroChIP-seq Kit',
'C01010132',
'680',
$('#CartQuantity').val());" name="keepshop" id="keepshop" type="submit">Keep shopping</button> </div>
</div>
</div>
</div>
</form><a class="close-reveal-modal" aria-label="Close">×</a></div><!-- END: ADD TO CART MODAL --><a href="#" id="true-microchip-kit-x16-16-rxns" data-reveal-id="cartModal-1856" class="" style="color:#B21329"><i class="fa fa-cart-plus"></i></a>
</div>
</div>
<div class="small-12 columns" >
<h6 style="height:60px">True MicroChIP-seq Kit</h6>
</div>
</div>
</li>
'
$related = array(
'id' => '1856',
'antibody_id' => null,
'name' => 'True MicroChIP-seq Kit',
'description' => '<p><a href="https://www.diagenode.com/files/products/kits/truemicrochipseq-kit-manual.pdf"><img src="https://www.diagenode.com/img/buttons/bt-manual.png" /></a></p>
<p>The <b>True </b><b>MicroChIP-seq</b><b> kit </b>provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as <b>10 000 cells</b>, including <b>FACS sorted cells</b>. The kit can be used for chromatin preparation for downstream ChIP-qPCR or ChIP-seq analysis. The <b>complete kit</b> contains everything you need for start-to-finish ChIP including all validated buffers and reagents for chromatin shearing, immunoprecipitation and DNA purification for exceptional <strong>ChIP-qPCR</strong> or <strong>ChIP-seq</strong> results. In addition, positive control antibodies and negative control PCR primers are included for your convenience and assurance of result sensitivity and specificity.</p>
<p>The True MicroChIP-seq kit offers unique benefits:</p>
<ul>
<li>An <b>optimized chromatin preparation </b>protocol compatible with low number of cells (<b>10.000</b>) in combination with the Bioruptor™ shearing device</li>
<li>Most <b>complete kit </b>available (covers all steps and includes control antibodies and primers)</li>
<li><b>Magnetic beads </b>make ChIP easy, fast, and more reproducible</li>
<li>MicroChIP DiaPure columns (included in the kit) enable the <b>maximum recovery </b>of immunoprecipitation DNA suitable for any downstream application</li>
<li><b>Excellent </b><b>ChIP</b><b>-seq </b>result when combined with <a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq">MicroPlex</a><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"> Library Preparation kit </a>adapted for low input</li>
</ul>
<p>For fast ChIP-seq on low input – check out Diagenode’s <a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">µ</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns">ChIPmentation</a><a href="https://www.diagenode.com/en/p/uchipmentation-for-histones-24-rxns"> for histones</a>.</p>
<p><sub>The True MicroChIP-seq kit, Cat. No. C01010132 is an upgraded version of the kit True MicroChIP, Cat. No. C01010130, with the new validated protocols (e.g. FACS sorted cells) and MicroChIP DiaPure columns included in the kit.</sub></p>',
'label1' => 'Characteristics',
'info1' => '<ul>
<li><b>Revolutionary:</b> Only 10,000 cells needed for complete ChIP-seq procedure</li>
<li><b>Validated on</b> studies for histone marks</li>
<li><b>Automated protocol </b>for the IP-Star<sup>®</sup> Compact Automated Platform available</li>
</ul>
<p></p>
<p>The True MicroChIP-seq kit protocol has been optimized for the use of 10,000 - 100,000 cells per immunoprecipitation reaction. Regarding chromatin immunoprecipitation, three protocol variants have been optimized:<br />starting with a batch, starting with an individual sample and starting with the FACS-sorted cells.</p>
<div><button id="readmorebtn" style="background-color: #b02736; color: white; border-radius: 5px; border: none; padding: 5px;">Show Workflow</button></div>
<p><br /> <img src="https://www.diagenode.com/img/product/kits/workflow-microchip.png" id="workflowchip" class="hidden" width="600px" /></p>
<p>
<script type="text/javascript">// <![CDATA[
const bouton = document.querySelector('#readmorebtn');
const workflow = document.getElementById('workflowchip');
bouton.addEventListener('click', () => workflow.classList.toggle('hidden'))
// ]]></script>
</p>
<div class="extra-spaced" align="center"></div>
<div class="row">
<div class="carrousel" style="background-position: center;">
<div class="container">
<div class="row" style="background: rgba(255,255,255,0.1);">
<div class="large-12 columns truemicro-slider" id="truemicro-slider">
<div>
<h3>High efficiency ChIP on 10,000 cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/true-micro-chip-histone-results.png" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 1. </strong>ChIP efficiency on 10,000 cells. ChIP was performed on human Hela cells using the Diagenode antibodies <a href="https://www.diagenode.com/en/p/h3k4me3-polyclonal-antibody-premium-50-ug-50-ul">H3K4me3</a> (Cat. No. C15410003), <a href="https://www.diagenode.com/en/p/h3k27ac-polyclonal-antibody-classic-50-mg-42-ml">H3K27ac</a> (C15410174), <a href="https://www.diagenode.com/en/p/h3k9me3-polyclonal-antibody-classic-50-ug">H3K9me3</a> (C15410056) and <a href="https://www.diagenode.com/en/p/h3k27me3-polyclonal-antibody-classic-50-mg-34-ml">H3K27me3</a> (C15410069). Sheared chromatin from 10,000 cells and 0.1 µg (H3K27ac), 0.25 µg (H3K4me3 and H3K27me3) or 0.5 µg (H3K9me3) of the antibody were used per IP. Corresponding amount of IgG was used as control. Quantitative PCR was performed with primers for corresponding positive and negative loci. Figure shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).</small></p>
</center></div>
</div>
<div>
<h3>True MicroChIP-seq protocol in a combination with MicroPlex library preparation kit results in reliable and accurate sequencing data</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig2-truemicro.jpg" alt="True MicroChip results" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 2.</strong> Integrative genomics viewer (IGV) visualization of ChIP-seq experiments using 50.000 of K562 cells. ChIP has been performed accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). The above figure shows the peaks from ChIP-seq experiments using the following antibodies: H3K4me1 (C15410194), H3K9/14ac (C15410200), H3K27ac (C15410196) and H3K36me3 (C15410192).</small></p>
</center></div>
</div>
<div>
<h3>Successful chromatin profiling from 10.000 of FACS-sorted cells</h3>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><img src="https://www.diagenode.com/img/product/kits/fig3ab-truemicro.jpg" alt="small non coding RNA" width="800px" /></div>
<div class="large-10 small-12 medium-10 large-centered medium-centered small-centered columns"><center>
<p><small><strong>Figure 3.</strong> (A) Integrative genomics viewer (IGV) visualization of ChIP-seq experiments and heatmap 3kb upstream and downstream of the TSS (B) for H3K4me3. ChIP has been performed using 10.000 of FACS-sorted cells (K562) and H3K4me3 antibody (C15410003) accordingly to True MicroChIP protocol followed by the library preparation using MicroPlex Library Preparation Kit (C05010001). Data were compared to ENCODE standards.</small></p>
</center></div>
</div>
</div>
</div>
</div>
</div>
</div>
<p>
<script type="text/javascript">// <![CDATA[
$('.truemicro-slider').slick({
arrows: true,
dots: true,
autoplay:true,
autoplaySpeed: 3000
});
// ]]></script>
</p>',
'label2' => 'Additional solutions compatible with the True MicroChIP-seq Kit',
'info2' => '<p><span style="font-weight: 400;">The <a href="https://www.diagenode.com/en/p/chromatin-shearing-optimization-kit-high-sds-100-million-cells">Chromatin EasyShear Kit – High SDS</a></span><span style="font-weight: 400;"> Recommended for the optimizing chromatin shearing.</span></p>
<p><a href="https://www.diagenode.com/en/categories/chip-seq-grade-antibodies"><span style="font-weight: 400;">ChIP-seq grade antibodies</span></a><span style="font-weight: 400;"> for high yields, specificity, and sensitivity.</span></p>
<p><span style="font-weight: 400;">Check the list of available </span><a href="https://www.diagenode.com/en/categories/primer-pairs"><span style="font-weight: 400;">primer pairs</span></a><span style="font-weight: 400;"> designed for high specificity to specific genomic regions.</span></p>
<p><span style="font-weight: 400;">For library preparation of immunoprecipitated samples we recommend to use the </span><b> </b><a href="https://www.diagenode.com/en/categories/library-preparation-for-ChIP-seq"><span style="font-weight: 400;">MicroPlex Library Preparation Kit</span></a><span style="font-weight: 400;"> - validated for library preparation from picogram inputs.</span></p>
<p><span style="font-weight: 400;">For IP-Star Automation users, check out the </span><a href="https://www.diagenode.com/en/p/auto-true-microchip-kit-16-rxns"><span style="font-weight: 400;">automated version</span></a><span style="font-weight: 400;"> of this kit.</span></p>
<p><span style="font-weight: 400;">Application note: </span><a href="https://www.diagenode.com/files/application_notes/Diagenode_AATI_Joint.pdf"><span style="font-weight: 400;">Best Workflow Practices for ChIP-seq Analysis with Small Samples</span></a></p>
<p></p>',
'label3' => 'Species, cell lines, tissues tested',
'info3' => '<p>The True MicroChIP-seq kit is compatible with a broad variety of cell lines, tissues and species - some examples are shown below. Other species / cell lines / tissues can be used with this kit.</p>
<p><strong>Cell lines:</strong></p>
<p>Bovine: blastocysts,<br />Drosophila: embryos, salivary glands<br />Human: EndoC-ẞH1 cells, HeLa cells, PBMC, urothelial cells<br />Mouse: adipocytes, B cells, blastocysts, pre-B cells, BMDM cells, chondrocytes, embryonic stem cells, KH2 cells, LSK cells, macrophages, MEP cells, microglia, NK cells, oocytes, pancreatic cells, P19Cl6 cells, RPE cells,</p>
<p>Other cell lines / species: compatible, not tested</p>
<p><strong>Tissues:</strong></p>
<p>Horse: adipose tissue</p>
<p>Mice: intestine tissue</p>
<p>Other tissues: not tested</p>',
'format' => '20 rxns',
'catalog_number' => 'C01010132',
'old_catalog_number' => 'C01010130',
'sf_code' => 'C01010132-',
'type' => 'RFR',
'search_order' => '04-undefined',
'price_EUR' => '625',
'price_USD' => '680',
'price_GBP' => '575',
'price_JPY' => '102405',
'price_CNY' => '',
'price_AUD' => '1700',
'country' => 'ALL',
'except_countries' => 'None',
'quote' => false,
'in_stock' => false,
'featured' => true,
'no_promo' => false,
'online' => true,
'master' => true,
'last_datasheet_update' => '0000-00-00',
'slug' => 'true-microchip-kit-x16-16-rxns',
'meta_title' => 'True MicroChIP-seq Kit | Diagenode C01010132',
'meta_keywords' => '',
'meta_description' => 'True MicroChIP-seq Kit provides a robust ChIP protocol suitable for the investigation of histone modifications within chromatin from as few as 10 000 cells, including FACS sorted cells. Compatible with ChIP-qPCR as well as ChIP-seq.',
'modified' => '2023-04-20 16:06:10',
'created' => '2015-06-29 14:08:20',
'ProductsRelated' => array(
'id' => '1743',
'product_id' => '1923',
'related_id' => '1856'
),
'Image' => array(
(int) 0 => array(
'id' => '1775',
'name' => 'product/kits/chip-kit-icon.png',
'alt' => 'ChIP kit icon',
'modified' => '2018-04-17 11:52:29',
'created' => '2018-03-15 15:50:34',
'ProductsImage' => array(
[maximum depth reached]
)
)
)
)
$rrbs_service = array(
(int) 0 => (int) 1894,
(int) 1 => (int) 1895
)
$chipseq_service = array(
(int) 0 => (int) 2683,
(int) 1 => (int) 1835,
(int) 2 => (int) 1836,
(int) 3 => (int) 2684,
(int) 4 => (int) 1838,
(int) 5 => (int) 1839,
(int) 6 => (int) 1856
)
$labelize = object(Closure) {
}
$old_catalog_number = ''
$country_code = 'US'
$label = '<img src="/img/banners/banner-customizer-back.png" alt=""/>'
$document = array(
'id' => '221',
'name' => 'MicroChIP DiaPure columns',
'description' => '<div class="page" title="Page 4">
<div class="section">
<div class="layoutArea">
<div class="column">
<ul>
<li>
<p><span>Quick (2 minute) recovery of ultra-pure DNA from chromatin immunoprecipitation (ChIP) assays, cell lysates, Proteinase K digested samples, PCRs and other enzymatic reactions. </span></p>
</li>
<li>
<p><span>Column design allows DNA to be eluted at high concentrations into minimal volumes (</span><span>≥ </span><span>6 μl) of water or low salt buffer. </span></p>
</li>
<li>
<p><span>Eluted DNA is ideal for PCR amplification, arrays, DNA quantification, Southern blot analysis, and other molecular applications. </span></p>
</li>
<li>
<p><span>Omits the use of organic solvents and the need for ethanol precipitation. </span></p>
</li>
</ul>
</div>
</div>
</div>
</div>',
'image_id' => null,
'type' => 'Manual',
'url' => 'files/products/reagents/MicroChIP_DiaPure_manual.pdf',
'slug' => 'microchip-diapure-manual',
'meta_keywords' => '',
'meta_description' => '',
'modified' => '2015-10-01 20:41:09',
'created' => '2015-07-07 11:47:43',
'ProductsDocument' => array(
'id' => '1227',
'product_id' => '1923',
'document_id' => '221'
)
)
$sds = array(
'id' => '175',
'name' => 'MicroChIP DiaPure columns SDS JP ja',
'language' => 'ja',
'url' => 'files/SDS/MicroChIP_DiaPure/SDS-C03040001-MicroChIP_DiaPure_columns-JP-ja-1_1.pdf',
'countries' => 'JP',
'modified' => '2020-06-08 15:18:52',
'created' => '2020-06-08 15:18:52',
'ProductsSafetySheet' => array(
'id' => '343',
'product_id' => '1923',
'safety_sheet_id' => '175'
)
)
$publication = array(
'id' => '2895',
'name' => 'Methionine-dependent histone methylation at developmentally important gene loci in mouse preimplantation embryos',
'authors' => 'Kudo M, Ikeda S, Sugimoto M, Kume S',
'description' => '<p>The involvement of specific nutrients in epigenetic gene regulation is a possible mechanism underlying nutrition-directed phenotypic alteration. However, the involvement of nutrients in gene-specific epigenetic regulation remains poorly understood. Methionine has been received attention as a possible nutrient involved in epigenetic modifications, as it is a precursor of the universal methyl donor for epigenetic methylation of DNA and histones. In the present study, the disruption of methionine metabolism by ethionine, an antimetabolite of methionine, induced abnormally higher expression of genes related to cell lineage differentiation and resulted in impaired blastocyst development of mouse preimplantation embryos in vitro. These effects were mitigated by the presence of methionine. Importantly, ethionine treatment induced lower trimethylation of histone H3 lysine 9 but did not affect methylation of DNA in the promoter regions of the examined genes. These results demonstrated that intact methionine metabolism is required for proper epigenetic histone modifications and normal expression of developmentally important genes during preimplantation development.</p>',
'date' => '2015-12-01',
'pmid' => 'http://www.ncbi.nlm.nih.gov/pubmed/26372092',
'doi' => '10.1016/j.jnutbio.2015.08.009',
'modified' => '2016-04-26 10:21:24',
'created' => '2016-04-26 10:21:24',
'ProductsPublication' => array(
'id' => '1173',
'product_id' => '1923',
'publication_id' => '2895'
)
)
$externalLink = ' <a href="http://www.ncbi.nlm.nih.gov/pubmed/26372092" target="_blank"><i class="fa fa-external-link"></i></a>'include - APP/View/Products/view.ctp, line 755
View::_evaluate() - CORE/Cake/View/View.php, line 971
View::_render() - CORE/Cake/View/View.php, line 933
View::render() - CORE/Cake/View/View.php, line 473
Controller::render() - CORE/Cake/Controller/Controller.php, line 963
ProductsController::slug() - APP/Controller/ProductsController.php, line 1052
ReflectionMethod::invokeArgs() - [internal], line ??
Controller::invokeAction() - CORE/Cake/Controller/Controller.php, line 491
Dispatcher::_invoke() - CORE/Cake/Routing/Dispatcher.php, line 193
Dispatcher::dispatch() - CORE/Cake/Routing/Dispatcher.php, line 167
[main] - APP/webroot/index.php, line 118
×