Monoclonal antibody raised in mouse against the amino-terminal domain of human TBP (TATA box binding protein).
TBP Antibody
Catalog Number
Format
Price
Other format

Alternative names: GTF2D, GTF2D1, SCA17, TF2D, TFIID
Monoclonal antibody raised in mouse against the amino-terminal domain of human TBP (TATA box binding protein).
| Lot | DA-0010 |
|---|---|
| Concentration | 8 µg/µl |
| Species reactivity | Human, mouse |
| Type | Monoclonal ChIP grade, ChIP-seq grade |
| Purity | Ammonium sulphate purified |
| Host | Mouse |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 4 - 5 μg/IP | Fig 1, 2 |
| WB | 1:500 | Fig 3 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-5 μl per IP.
- Validation Data
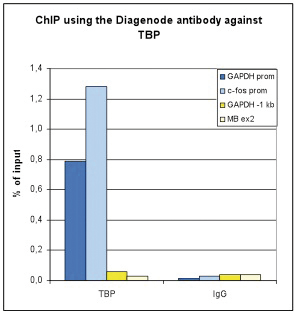
Figure 1 ChIP-seq results obtained with the Diagenode monoclonal antibody directed against TBP
ChIP was performed with 5 μg of the Diagenode antibody against TBP (Cat. No. C15200002) on sheared chromatin from 1 million HeLaS3 cells using the “Auto Histone ChIP-seq” kit (Cat. No. C01010022) on the IP-Star automated system. The IP’d DNA was analysed by QPCR with optimized PCR primer pairs for the promoters of the active GAPDH and c-fos genes, used as positive control targets, and for a region 1 kb upstream of the GAPDH promoter and the coding region of the inactive MB gene, used as negative control targets (figure 2A). The IP’d DNA was subsequently analysed with an Illumina Genome Analyzer. Library preparation, cluster generation and sequencing were performed according to the manufacturer’s instructions. The 36 bp tags were aligned to the human genome using the ELAND algorithm. Figure 2 shows the peak distribution in 50 kb regions surrounding the GAPDH, c-fos, ACTB and MCL1 genes (figure 2B, C, D and E, respectively). These results clearly show a localisation of TBP at the promoters of actively transcribed genes.A.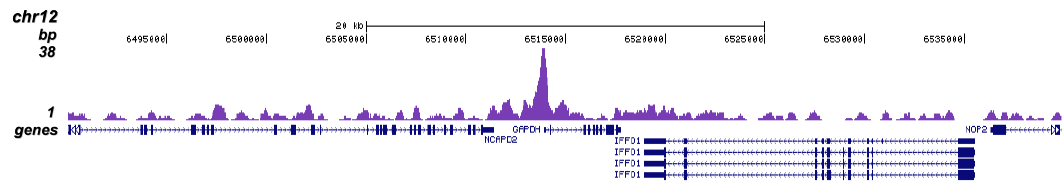
B.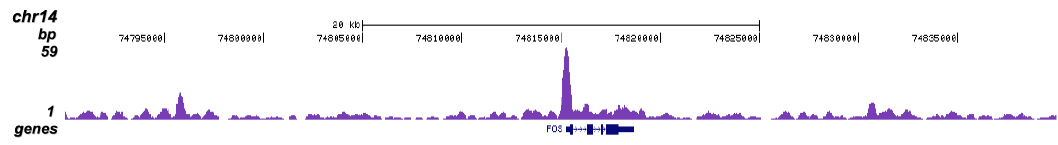
C.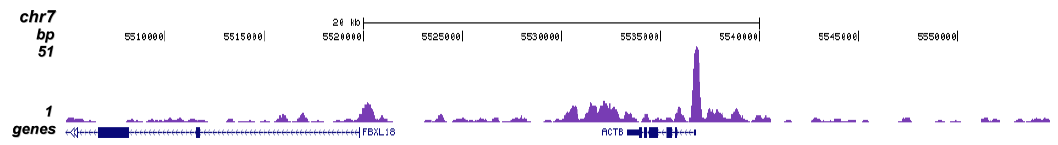
D.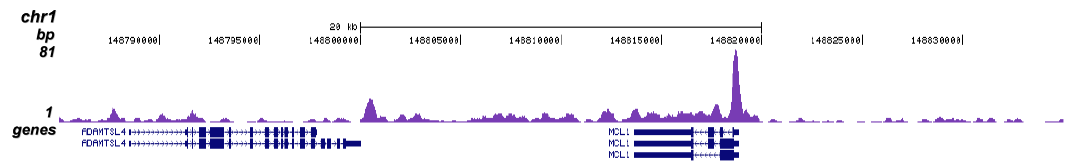
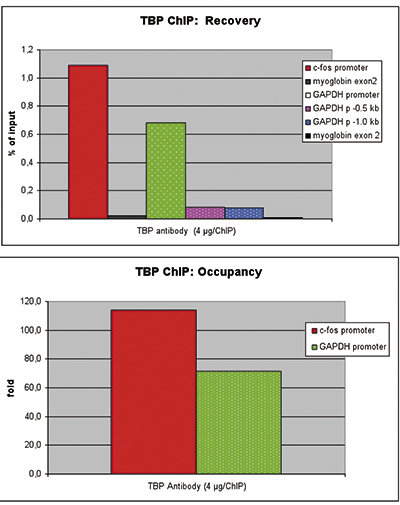
Figure 2 ChIP results obtained with the Diagenode monoclonal antibody against TBP
ChIP assays were performed using U2OS cells, the Diagenode antibody directed against TBP (Cat. No. C15200002) and optimized primer sets for qPCR. Sheared chromatin from 1x10e6 cells and 4 μg of antibody were used per ChIP experiment. QPCR was performed with primers for the promoter of the c-fos and GAPDH genes (Cat. No. C17011004 and C17011001), a region 0.5 and 1 kb upstream of the GAPDH promoter (Cat. No. C17011002 and C17011003), respectively, and for exon 2 of the myoglobin gene (cat. No. C17011006) as a negative control. Figure 1 shows the recovery (the relative amount of immunoprecipitated DNA compared to input DNA) and the occupancy (ratio +/- control target). These results demonstrate the occupancy of both promoters by TBP.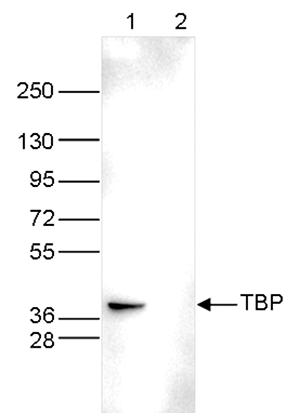
Figure 3. Western blot analysis using the Diagenode monoclonal antibody directed against TBP
Whole cell extracts (40 μg) from HeLa cells transfected with TBP siRNA (lane 2) and from an untransfected control (lane 1) were analysed by Western blot using the Diagenode antibody against TBP (Cat. No. C15200002) diluted 1:500 in TBSTween containing 5% skimmed milk. The position of the protein of interest is indicated on the right; the marker (in kDa) is shown on the left. - Publications

How to properly cite our product/service in your work
We strongly recommend using this: TBP Antibody (Hologic Diagenode Cat# C15200002 Lot# DA-0010 ). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
Enhanced frequency of transcription pre-initiation complexes assemblyafter exposure to UV irradiation results in increased repair activity andreduced probabilities for mutagenesis.
Liakos A. et al.
In addition to being essential for gene expression, transcription is crucial for the maintenance of genome integrity. Here, we undertook a systematic approach, to monitor the assembly kinetics of the pre-initiating RNA Polymerase (Pol) II at promoters at steady state and different stages during recovery from UV irra...A leukemia-protective germline variant mediates chromatin moduleformation via transcription factor nucleation.
Llimos G. et al.
Non-coding variants coordinate transcription factor (TF) binding and chromatin mark enrichment changes over regions spanning >100 kb. These molecularly coordinated regions are named "variable chromatin modules" (VCMs), providing a conceptual framework of how regulatory variation might shape complex traits....R-Loops Promote Antisense Transcription across the Mammalian Genome
Tan-Wong Sue Mei, Dhir Somdutta, Proudfoot Nick J.
Widespread antisense long noncoding RNA (lncRNA) overlap with many protein-coding genes in mammals and emanate from gene promoter, enhancer, and termination regions. However, their origin and biological purpose remain unclear. We show that these antisense lncRNA can be generated by R-loops that form when nascent tra...Adenovirus E1A Activation Domain Regulates H3 Acetylation Affecting Varied Steps in Transcription at Different Viral Promoters.
Hsu E, Pennella MA, Zemke NR, Eng C, Berk AJ
How histone acetylation promotes transcription is not clearly understood. Here, we confirm an interaction between p300 and the adenovirus 2 large E1A activation domain (AD) and map the interacting regions in E1A by observing colocalization at an integrated array of fusions of LacI-mCherry to E1A fragments with YFP-p...Condensin targets and reduces unwound DNA structures associated with transcription in mitotic chromosome condensation
Sutani T, Sakata T, Nakato R, Masuda K, Ishibashi M, Yamashita D, Suzuki Y, Hirano T, Bando M, Shirahige K
Chromosome condensation is a hallmark of mitosis in eukaryotes and is a prerequisite for faithful segregation of genetic material to daughter cells. Here we show that condensin, which is essential for assembling condensed chromosomes, helps to preclude the detrimental effects of gene transcription on mitotic condens...A pro-apoptotic function of iASPP by stabilizing p300 and CBP through inhibition of BRMS1 E3 ubiquitin ligase activity.
Kramer D, Schön M, Bayerlová M, Bleckmann A, Schön MP, Zörnig M, Dobbelstein M
The p53 family and its cofactors are potent inducers of apoptosis and form a barrier to cancer. Here, we investigated the impact of the supposedly inhibitory member of the apoptosis-stimulating protein of p53, iASPP, on the activity of the p53 homolog TAp73, and its cofactors p300 and CBP. We found that iASPP intera...The unfolded protein response and the phosphorylations of activating transcription factor 2 in the trans-activation of il23a promoter produced by β-glucans.
Rodríguez M, Domingo E, Alonso S, Frade JG, Eiros J, Crespo MS, Fernández N
Current views on the control of IL-23 production focus on the regulation of il23a, the gene encoding IL-23 p19, by NF-κB in combination with other transcription factors. C/EBP homologous protein (CHOP), X2-Box-binding protein 1 (XBP1), activator protein 1 (AP1), SMAD, CCAAT/enhancer-binding protein (C/EBPβ), and cAM...ERG and FLI1 binding sites demarcate targets for aberrant epigenetic regulation by AML1-ETO in acute myeloid leukemia.
Martens JH, Mandoli A, Simmer F, Wierenga BJ, Saeed S, Singh AA, Altucci L, Vellenga E, Stunnenberg HG
ERG and FLI1 are closely related members of the ETS family of transcription factors and have been identified as essential factors for the function and maintenance of normal hematopoietic stem cells. Here, genome-wide analysis revealed that both ERG and FLI1 occupy similar genomic regions as AML1-ETO in t(8;21) AMLs ...Recruitment of histone deacetylase 3 to the interferon-a gene promoters attenuates interferon expression.
Génin P, Lin R, Hiscott J, Civas A
BACKGROUND: Induction of Type I Interferon (IFN) genes constitutes an essential step leading to innate immune responses during virus infection. Sendai virus (SeV) infection of B lymphoid Namalwa cells transiently induces the transcriptional expression of multiple IFN-A genes. Although transcriptional activation of I...Coactivation of GR and NFKB alters the repertoire of their binding sites and target genes.
Rao NA, McCalman MT, Moulos P, Francoijs KJ, Chatziioannou A, Kolisis FN, Alexis MN, Mitsiou DJ, Stunnenberg HG
Glucocorticoid receptor (GR) exerts anti-inflammatory action in part by antagonizing proinflammatory transcription factors such as the nuclear factor kappa-b (NFKB). Here, we assess the crosstalk of activated GR and RELA (p65, major NFKB component) by global identification of their binding sites and target genes. We...Transcription initiation platforms and GTF recruitment at tissue-specific enhancers and promoters.
Koch F, Fenouil R, Gut M, Cauchy P, Albert TK, Zacarias-Cabeza J, Spicuglia S, de la Chapelle AL, Heidemann M, Hintermair C, Eick D, Gut I, Ferrier P, Andrau JC
Recent work has shown that RNA polymerase (Pol) II can be recruited to and transcribe distal regulatory regions. Here we analyzed transcription initiation and elongation through genome-wide localization of Pol II, general transcription factors (GTFs) and active chromatin in developing T cells. We show that Pol II an...Role of p53 serine 46 in p53 target gene regulation.
Smeenk L, van Heeringen SJ, Koeppel M, Gilbert B, Janssen-Megens E, Stunnenberg HG, Lohrum M
The tumor suppressor p53 plays a crucial role in cellular growth control inducing a plethora of different response pathways. The molecular mechanisms that discriminate between the distinct p53-responses have remained largely elusive. Here, we have analyzed the p53-regulated pathways induced by Actinomycin D and Etop...Control of the differentiation of regulatory T cells and T(H)17 cells by the DNA-binding inhibitor Id3.
Maruyama T, Li J, Vaque JP, Konkel JE, Wang W, Zhang B, Zhang P, Zamarron BF, Yu D, Wu Y, Zhuang Y, Gutkind JS, Chen W
The molecular mechanisms that direct transcription of the gene encoding the transcription factor Foxp3 in CD4(+) T cells remain ill-defined. We show here that deletion of the DNA-binding inhibitor Id3 resulted in the defective generation of Foxp3(+) regulatory T cells (T(reg) cells). We identify two transforming gro...High-resolution analysis of epigenetic changes associated with X inactivation.
Marks H, Chow JC, Denissov S, Françoijs KJ, Brockdorff N, Heard E, Stunnenberg HG
Differentiation of female murine ES cells triggers silencing of one X chromosome through X-chromosome inactivation (XCI). Immunofluorescence studies showed that soon after Xist RNA coating the inactive X (Xi) undergoes many heterochromatic changes, including the acquisition of H3K27me3. However, the mechanisms that ...TIPT2 and geminin interact with basal transcription factors to synergize in transcriptional regulation.
Pitulescu ME, Teichmann M, Luo L, Kessel M
BACKGROUND: The re-replication inhibitor Geminin binds to several transcription factors including homeodomain proteins, and to members of the polycomb and the SWI/SNF complexes. RESULTS: Here we describe the TATA-binding protein-like factor-interacting protein (TIPT) isoform 2, as a strong binding partner of Geminin...Identification of novel functional TBP-binding sites and general factor repertoires
Denissov S, van Driel M, Voit R, Hekkelman M, Hulsen T, Hernandez N, Grummt I, Wehrens R, Stunnenberg H.
Our current knowledge of the general factor requirement in transcription by the three mammalian RNA polymerases is based on a small number of model promoters. Here, we present a comprehensive chromatin immunoprecipitation (ChIP)-on-chip analysis for 28 transcription factors on a large set of known and novel TATA-bin...