HDAC1 (UniProt/Swiss-Prot entry Q13547) catalyses the deacetylation of lysine residues on the N-terminal part of the core histones (H2A, H2B, H3 and H4). Acetylation and deacetylation of these highly conserved lysine residues is important for the control of gene expression and HDAC activity is often associated with gene repression. Histone deacetylation is established by the formation of large multiprotein complexes. HDAC1 also interacts with the retinoblastoma tumor suppressor protein and is able to deacetylate p53. Therefore, it also plays an essential role in cell proliferation and differentiation and in apoptosi.
HDAC1 Antibody - ChIP-seq Grade

Alternative names: HD1, RPD3, RPD3L1, GON-10
Polyclonal antibody raised in rabbit against the C-terminal region of human HDAC1 (Histone deacetylase 1), using a KLH-conjugated synthetic peptide.
| Lot | A21-001P |
|---|---|
| Concentration | 1.73 μg/μl |
| Species reactivity | Human, mouse |
| Type | Polyclonal |
| Purity | Affinity purified |
| Host | Rabbit |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 2 μg/ChIP | Fig 1, 2 |
| ELISA | 1:4,000 | Fig 3 |
| Western Blotting | 1:1,000 | Fig 4, 5 |
| Protein array | 1:100,000 | Fig 6 |
| Immunofluorescence | 1:500 | Fig 7 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-5 μg per IP.
- Validation Data
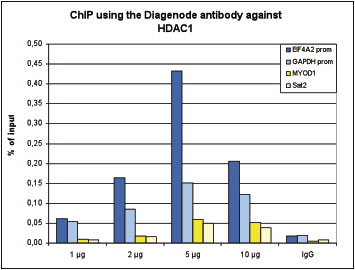
Figure 1. ChIP results obtained with the Diagenode antibody directed against HDAC1
ChIP was performed with the Diagenode antibody against HDAC1 (Cat. No. C15410325) on sheared chromatin from 4,000,000 HeLa cells. An antibody titration consisting of 1, 2, 5 and 10 μg per ChIP experiment was analysed. IgG (2 μg/IP) was used as negative IP control. QPCR was performed with primers specific for the EIF4A2 and GAPDH promoters, used as positive controls, and for the MYOD1 gene and Sat2 satellite repeat, used as negative controls. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).
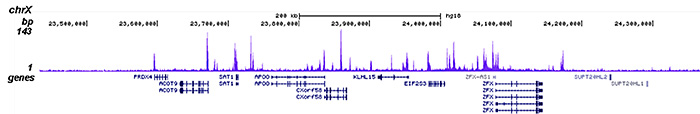
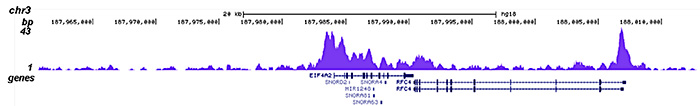
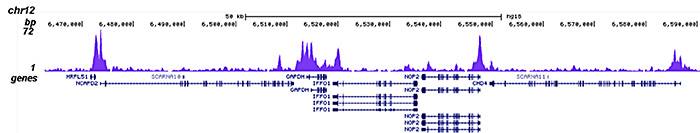
Figure 2. ChIP-seq results obtained with the Diagenode antibody directed against HDAC1
ChIP was performed on sheared chromatin from 4,000,000 HeLa cells using 2 μg of the Diagenode antibody against HDAC1 (Cat. No. C15410325) as described above. The IP’d DNA was subsequently analysed on an Illumina HiSeq 2000. Library preparation, cluster generation and sequencing were performed according to the manufacturer’s instructions. The 50 bp tags were aligned to the human genome using the BWA algorithm. Figure 2 shows the peak distribution along the complete sequence and a 1 Mb region of the X-chromosome (figure 2A and B) and in two regions surrounding the GAPDH and EIF4A2 positive control genes, respectively (figure 2C and D)..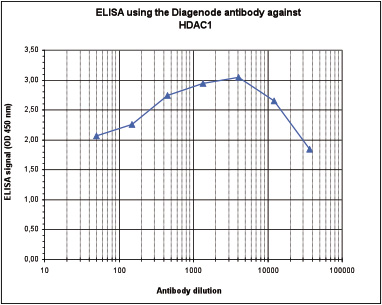
Figure 3. Determination of the antibody titer
To determine the titer of the antibody, an ELISA was performed using a serial dilution of Diagenode antibody directed against HDAC1 (Cat. No. C15410325), crude serum and flow through. The plates were coated with the peptide used for immunization of the rabbit. By plotting the absorbance against the antibody dilution (Figure 2), the titer of the antibody was estimated to be 1:75,000.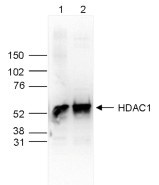
Figure 4. Western blot analysis using the Diagenode antibody directed against HDAC1
Whole cell extracts (25 μg, lane 1) and nuclear extracts (25 μg, lane 2) from HeLa cells were analysed by Western blot using the Diagenode antibody against HDAC1 (Cat. No. C15410325) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right (expected size: 55 kDa); the marker (in kDa) is shown on the left.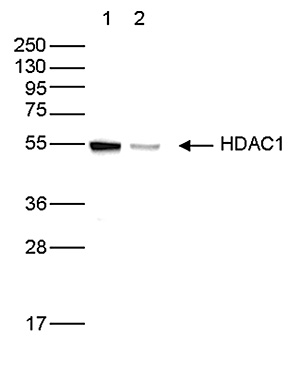
Figure 5. Western blot analysis using the Diagenode antibody directed against HDAC1
Whole cell extracts (50 µg) from HeLa cells transfected with HDAC1 siRNA (lane 2) and from an untransfected control (lane 1) were analysed by Western blot using the Diagenode antibody against HDAC1 (Cat. No. C15410325) diluted 1:1,000 in TBS-Tween containing 5% skimmed milk. The position of the protein of interest is indicated on the right (expected size: 55 kDa); the marker (in kDa) is shown on the left.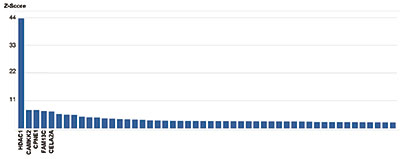
Figure 6. Protein array analysis with the Diagenode antibody directed against HDAC1
The specificity of the Diagenode antibody against HDAC1 (Cat. No. C15410325) was demonstrated using the HuProt human protein microarray (CDI Laboratories), a protein array containing more than 19,000 human proteins. The antibody was used at a dilution of 1:100,000. Figure 6 shows the Z-score of the signal intensity (mean value of the duplicate spots on the array). The names of the proteins with 5 highest Z-scores are indicated at the bottom. This figure clearly shows the high specificity of the antibody for HDAC1.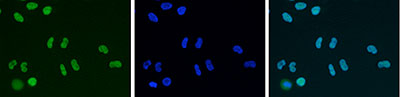
Figure 7. Immunofluorescence using the Diagenode antibody directed against HDAC1
HeLa cells were stained with the Diagenode antibody against HDAC1 (Cat. No. C15410325) and with DAPI. Cells were fixed with 4% formaldehyde for 10’ and blocked with PBS/TX-100 containing 5% normal goat serum and 1% BSA. The cells were immunofluorescently labelled with the HDAC1 antibody (left) diluted 1:500 in blocking solution followed by an anti-rabbit antibody conjugated to Alexa488. The middle panel shows staining of the nuclei with DAPI. A merge of the two stainings is shown on the right. - 出版物

How to properly cite our product/service in your work
We strongly recommend using this: HDAC1 Antibody - ChIP-seq Grade (Hologic Diagenode Cat# C15410325 Lot# A21-001P). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
NUP98 and RAE1 sustain progenitor function through HDAC-dependentchromatin targeting to escape from nucleolar localization.
Neely Amy E. et al.
Self-renewing somatic tissues rely on progenitors to support the continuous tissue regeneration. The gene regulatory network maintaining progenitor function remains incompletely understood. Here we show that NUP98 and RAE1 are highly expressed in epidermal progenitors, forming a separate complex in the nucleoplasm. ...Histone Deacetylases 1 and 2 target gene regulatory networks of nephronprogenitors to control nephrogenesis.
Liu Hongbing et al.
Our studies demonstrated the critical role of Histone deacetylases (HDACs) in the regulation of nephrogenesis. To better understand the key pathways regulated by HDAC1/2 in early nephrogenesis, we performed chromatin immunoprecipitation sequencing (ChIP-Seq) of Hdac1/2 on isolated nephron progenitor cells (NPCs) fro...Threonine phosphorylation of IκBζ mediates inhibition of selective proinflammatory target genes.
Grondona P, Bucher P, Schmitt A, Schönfeld C, Streibl B, Müller A, Essmann F, Liberatori S, Mohammed S, Hennig A, Kramer D, Schulze-Osthoff K, Hailfinger S
Transcription factors of the NF-κB family play a crucial role for immune responses by activating the expression of chemokines, cytokines and antimicrobial peptides involved in pathogen clearance. IκBζ, an atypical nuclear IκB protein and selective coactivator of particular NF-κB target g...Immunity drives regulation in cancer through NF-κB.
Collignon E, Canale A, Al Wardi C, Bizet M, Calonne E, Dedeurwaerder S, Garaud S, Naveaux C, Barham W, Wilson A, Bouchat S, Hubert P, Van Lint C, Yull F, Sotiriou C, Willard-Gallo K, Noel A, Fuks F
Ten-eleven translocation enzymes (TET1, TET2, and TET3), which induce DNA demethylation and gene regulation by converting 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), are often down-regulated in cancer. We uncover, in basal-like breast cancer (BLBC), genome-wide 5hmC changes related to regulation. We fu...Immunity drives TET1 regulation in cancer through NF-kB
Collignon E, Canale A, Al Wardi C, Bizet M, Calonne E, Dedeurwaerder S, Garaud S, Naveaux C, Barham W, Wilson A, Bouchat S, Hubert P, Van Lint C, Yull F, Sotiriou C, Willard-Gallo K, Noel A, Fuks F
Ten-eleven translocation enzymes (TET1, TET2, and TET3), which induce DNA demethylation and gene regulation by converting 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), are often down-regulated in cancer. We uncover, in basal-like breast cancer (BLBC), genome-wide 5hmC changes related to regulation. We fu...Krüppel-like transcription factor KLF10 suppresses TGFβ-induced epithelial-to-mesenchymal transition via a negative feedback mechanism
Mishra V.K. et al.
TGFβ-SMAD signaling exerts a contextual effect that suppresses malignant growth early in epithelial tumorigenesis but promotes metastasis at later stages. Longstanding challenges in resolving this functional dichotomy may uncover new strategies to treat advanced carcinomas. The Krüppel-like transcription f...HDAC1 negatively regulates Bdnf and Pvalb required for parvalbumin interneuron maturation in an experience-dependent manner
Koh DX and Sng JC
During early postnatal development, neuronal circuits are sculpted by sensory experience provided by the external environment. This experience-dependent regulation of circuitry development consolidates the balance of excitatory-inhibitory (E/I) neurons in the brain. The cortical barrel-column that innervates a singl...Genome-wide hydroxymethylcytosine pattern changes in response to oxidative stress
Delatte B, Jeschke J, Defrance M, Bachman M, Creppe C, Calonne E, Bizet M, Deplus R, Marroquí L, Libin M, Ravichandran M, Mascart F, Eizirik DL, Murrell A, Jurkowski TP, Fuks F
The TET enzymes convert methylcytosine to the newly discovered base hydroxymethylcytosine. While recent reports suggest that TETs may play a role in response to oxidative stress, this role remains uncertain, and results lack in vivo models. Here we show a global decrease of hydroxymethylcytosine in cells treated w...SNAIL1 combines competitive displacement of ASCL2 and epigenetic mechanisms to rapidly silence the EPHB3 tumor suppressor in colorectal cancer.
Rönsch K, Jägle S, Rose K, Seidl M, Baumgartner F, Freihen V, Yousaf A, Metzger E, Lassmann S, Schüle R, Zeiser R, Michoel T, Hecht A
EPHB3 is a critical cellular guidance factor in the intestinal epithelium and an important tumor suppressor in colorectal cancer (CRC) whose expression is frequently lost at the adenoma-carcinoma transition when tumor cells become invasive. The molecular mechanisms underlying EPHB3 silencing are incompletely underst...Citrullination of DNMT3A by PADI4 regulates its stability and controls DNA methylation.
Deplus R, Denis H, Putmans P, Calonne E, Fourrez M, Yamamoto K, Suzuki A, Fuks F
DNA methylation is a central epigenetic modification in mammals, with essential roles in development and disease. De novo DNA methyltransferases establish DNA methylation patterns in specific regions within the genome by mechanisms that remain poorly understood. Here we show that protein citrullination by peptidylar...Dimethyl fumarate regulates histone deacetylase expression in astrocytes.
Kalinin S, Polak PE, Lin SX, Braun D, Guizzetti M, Zhang X, Rubinstein I, Feinstein DL
We previously showed that dimethyl fumarate (DMF) reduces inflammatory activation in astrocytes, involving activation of transcription factor Nrf2. However, the pathways causing Nrf2 activation were not examined. We now show that DMF modifies expression of histone deacetylases (HDACs) in primary rat astrocytes. Afte...Phosphorylation of p65(RelA) on Ser547 by ATM Represses NF-κB-Dependent Transcription of Specific Genes after Genotoxic Stress
Sabatel H, Di Valentin E, Gloire G, Dequiedt F, Piette J, Habraken YThe histone demethylase Kdm3a is essential to progression through differentiation.
Herzog M, Josseaux E, Dedeurwaerder S, Calonne E, Volkmar M, Fuks F
Histone demethylation has important roles in regulating gene expression and forms part of the epigenetic memory system that regulates cell fate and identity by still poorly understood mechanisms. Here, we examined the role of histone demethylase Kdm3a during cell differentiation, showing that Kdm3a is essential for ...HDAC1 Regulates Fear Extinction in Mice.
Bahari-Javan S, Maddalena A, Kerimoglu C, Wittnam J, Held T, Bähr M, Burkhardt S, Delalle I, Kügler S, Fischer A, Sananbenesi F
Histone acetylation has been implicated with the pathogenesis of neuropsychiatric disorders and targeting histone deacetylases (HDACs) using HDAC inhibitors was shown to be neuroprotective and to initiate neuroregenerative processes. However, little is known about the role of individual HDAC proteins during the path...Enhancer of Zeste 2 (EZH2) is up-regulated in malignant gliomas and in glioma stem-like cells.
Orzan F, Pellegatta S, Poliani PL, Pisati F, Caldera V, Menghi F, Kapetis D, Marras C, Schiffer D, Finocchiaro G
AIMS: Proteins of the Polycomb repressive complex 2 (PRC2) are epigenetic gene silencers and are involved in tumour development. Their oncogenic function might be associated with their role in stem cell maintenance. The histone methyltransferase Enhancer of Zeste 2 (EZH2) is a key member of PRC2 function: we have in...The core binding factor CBF negatively regulates skeletal muscle terminal differentiation.
Philipot O, Joliot V, Ait-Mohamed O, Pellentz C, Robin P, Fritsch L, Ait-Si-Ali S
BACKGROUND: Core Binding Factor or CBF is a transcription factor composed of two subunits, Runx1/AML-1 and CBF beta or CBFbeta. CBF was originally described as a regulator of hematopoiesis. METHODOLOGY/PRINCIPAL FINDINGS: Here we show that CBF is involved in the control of skeletal muscle terminal differentiation. I...Functional connection between deimination and deacetylation of histones.
Denis H, Deplus R, Putmans P, Yamada M, Métivier R, Fuks F
Histone methylation plays key roles in regulating chromatin structure and function. The recent identification of enzymes that antagonize or remove histone methylation offers new opportunities to appreciate histone methylation plasticity in the regulation of epigenetic pathways. Peptidylarginine deiminase 4 (PADI4; a...